脳梗塞後のニューロン再生過程の解析
名古屋市立大学大学院医学研究科脳神経科学研究所神経発達・再生医学分野
発行日:2019年12月30日Published: December 30, 2019
© 2019 日本神経化学会© 2019 The Japanese Society for Neurochemistry
哺乳類の脳では、ニューロンの大部分が胎生後期に神経幹細胞によって産生される。ニューロンは発達中の脳内を最終的な位置まで移動したのち成熟して、軸索や樹状突起を伸ばして神経回路を形成する。ニューロンの産生に続いて、グリアを産生した神経幹細胞は、自身もグリア細胞へ分化する。よって、脳卒中や外傷、神経変性疾患などでニューロンが死滅しても、新たなニューロンを産生して損傷を修復することはできず、永続的な神経機能の障害が生じる。しかし近年の研究によって、成熟した脳でも特定の領域では神経幹細胞が維持されて、ニューロンが持続的に産生されていることが明らかになった。「脳室下帯」と呼ばれる側脳室周囲の細胞層は、この現象がみられる成体脳内で最大の領域である1, 2)。
脳室下帯で産生された幼若なニューロンは、短い突起を前後に伸ばしたシンプルな形態で、脳の前端に位置する嗅球まで、成熟脳内を長距離にわたって高速で移動する3)。嗅球で介在ニューロンに分化して神経回路に組み込まれ、嗅覚系の神経回路において臭いの弁別や学習に関与する。また、脳梗塞や神経変性疾患でニューロンが脱落すると、新生ニューロンは傷害部に移動して成熟することが示されている4–6)。傷害部に供給される新生ニューロンを増やすことで神経機能の回復も促進される可能性が示唆されているが3, 7)、傷害部に移動した新生ニューロンの機能は分かっていない。しかし、この現象は哺乳類の脳内にもニューロン再生システムが潜在することを示唆しており、成熟脳内でのニューロン新生・再生のメカニズムを理解し、傷害部で効率よくニューロンを再生することが可能になれば、脳疾患の治療への応用も期待できる。本稿では、成体脳内での新生ニューロンの移動と脳梗塞後のニューロン再生メカニズムに関するこれまでの知見と、我々の研究成果を紹介する。
ニューロンの移動は胎生期に大規模に行われており、その制御メカニズムに関してこれまでに多くの知見が集積されてきた8–10)。細胞体の前方に伸びる先導突起には、一方向に配向した微小管が存在し、先端部はアクチンの重合が活発な成長円錐様の構造になっている。微小管・アクチンの動態や足場との接着の制御、誘引因子や反発因子の分布など、複雑な制御機構によってニューロンの移動方向や停止位置は精緻にコントロールされている。これらのメカニズムの一部は、成体脳内の新生ニューロンの移動制御にも関与している。一方、発達過程の脳とは異なり、神経突起が発達し多数のグリア細胞が存在する成体脳内を新生ニューロンが長距離移動するには、独自の移動制御メカニズムも必要である。
成体脳内では、脳室下帯で産生された新生ニューロンは、吻側移動流(RMS, rostral migratory stream)を経て嗅球へ移動する11)(図1左)。この間、新生ニューロンは細長い細胞塊を形成し、その中で互いを足場として移動・停止を交互に繰り返しながら前進する「鎖状細胞移動(chain migration)」と呼ばれる独自の様式で移動する。この鎖の中の新生ニューロンは、複数の小さな接着結合と非接着部を介して周囲の新生ニューロンと接しており、細胞接着因子12–15)や細胞骨格制御因子などが鎖状細胞塊の形成に関与することが分かっている。また、複数の誘引因子・反発因子が新生ニューロンの移動方向を制御している。
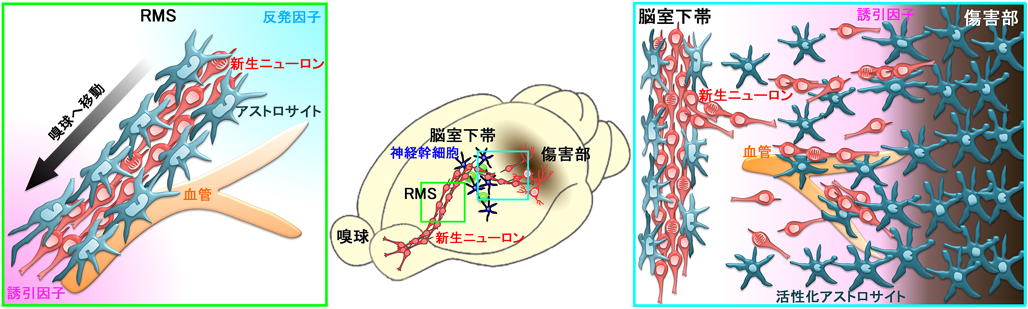
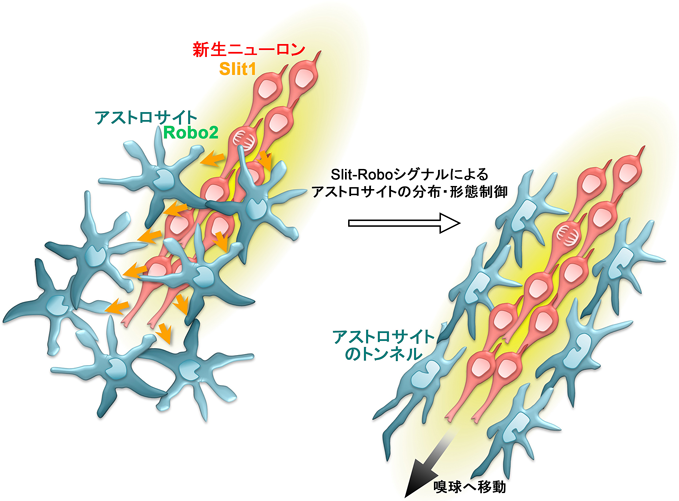
RMSでは、新生ニューロンの細胞塊は主要なグリア細胞であるアストロサイトがつくるトンネルによって被覆されている11)。この構造は、アストロサイトが未発達な幼若な脳では見られず、成体脳に特有のものである。RMSにおける新生ニューロンの一部は血管に沿って移動するが16, 17)、このときも新生ニューロンが直接に接触するのは血管ではなく、血管を取り巻くアストロサイトである。RMSのアストロサイトはトンネル形成によって新生ニューロンの周囲組織との相互作用を制御し、新生ニューロンの移動を制御する因子の分泌や濃度調節を行っている17–19)。我々は、移動している新生ニューロンがSlit-Roboシグナルを介して、このトンネルの形成・維持に関与していることを明らかにした20)(図2)。Slit1は細胞外に分泌され、膜貫通型受容体であるRoboに結合し、様々な組織の発生過程において、細胞の移動や突起伸長の方向を反発性に制御するガイダンス因子として機能している。脳室下帯で産生される新生ニューロンは、RMSを移動中にSlit1を発現していることが報告された21)。我々は、トンネルを形成するRMSのアストロサイトはその受容体Robo2を強く発現していることを見出し、新生ニューロン由来のSlit1が、Robo2を介してアストロサイトの分布や形態を変化させ、トンネルの形成・維持に寄与していることを示した。このトンネルは、新生ニューロンが成体脳内を高速で移動するのに必要であることから、新生ニューロンが自身の高速移動に必要な環境を整備するという、新規の移動制御メカニズムの存在が明らかになった。
生理的条件下では、脳室下帯で産生された新生ニューロンは上述のように嗅球のみに供給される。一方、脳梗塞や外傷、神経変性疾患など、多数のニューロンが脱落する様々な病態下では、脳室下帯で産生された新生ニューロンの一部は傷害部周囲に移動して成熟する4–6)(図1右)。この過程は、脳梗塞モデル動物において最もよく研究されている。脳梗塞で傷害された組織で産生されるケモカインや栄養因子・増殖因子などが、新生ニューロンを脳室下帯から傷害部へ誘引している22–25)。神経幹細胞を活性化したり、新生ニューロンの移動・生存を促進したりする様々な介入法によって、傷害後の神経機能の回復が促進されることが報告されているが、病態下での新生ニューロンの挙動や機能には、未だ不明な点が多い。
新生ニューロンの移動の研究には、臨床的に脳梗塞の責任動脈となることが多い中大脳動脈の起始部を一過性に閉塞して作成した、脳梗塞モデルマウスがよく用いられる26, 27)。このモデルでは、線条体の外側と隣接する側頭皮質に、ニューロンが完全に脱落した梗塞巣が形成させる。これに反応して、脳内では速やかに各種グリア細胞の増生が始まる。脳内の主要なグリア細胞であるアストロサイトは、梗塞巣とその周囲の広範な領域で活性化し、個々の細胞体・突起が肥大するとともに分裂して増生する28)。一方、線条体への新生ニューロンの移動は、脳梗塞1週後くらいから始まるため6, 29)、新生ニューロンは活性化アストロサイトが密集する領域を通過しなければ梗塞巣に近づくことができない(図1右)。しかし、活性化アストロサイトが新生ニューロンの移動にどのような影響を与えるかは分かっていなかった。
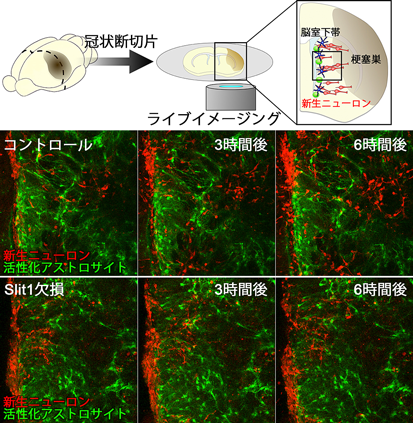
アストロサイトは細胞間隙を埋めるように不規則で複雑な微細突起を表面に形成しており、光学顕微鏡でこの形態を完全に描出することは難しい。そこで我々は、大野伸彦先生(自治医科大学)にご協力頂き、連続断層走査顕微鏡(serial block-face scanning electron microscope, SBF-SEM)を用いて脳梗塞巣に移動する新生ニューロンと活性化アストロサイトの空間的関係を詳しく調べた。活性化アストロサイトは非常に広範囲に突起を伸展させており、これらの突起が新生ニューロンの細胞塊の表面を覆うように接触していた(図3左)。また、梗塞後の脳スライスのライブイメージングから、活性化アストロサイトが密生する領域では、新生ニューロンの移動速度が低下して方向性も不安定になることが分かった。

梗塞後の線条体を移動する新生ニューロン、活性化アストロサイトにもSlit1, Robo2の発現が見られた。Slit1を欠損した新生ニューロンの活性化アストロサイト密集領域での移動は著しく抑制され(図4)、レンチウィルスベクターを用いて局所的に活性化アストロサイトのRobo2発現を抑制すると、その領域で新生ニューロンは移動を停止した。これらの結果から、活性化アストロサイトが分布する領域での新生ニューロンのスムーズな移動にSlit-Roboシグナルが必要であることが分かった30)。
SBF-SEMで観察した梗塞脳内の活性化アストロサイトの表面には、細かい突起が数多く存在するが、新生ニューロンと接する面は平滑で突起が存在しない(図3右)。そこで、新生ニューロン–活性化アストロサイト共培養実験系を構築し、重合アクチン可視化プローブを用いて31, 32)、新生ニューロンの接触によるアストロサイトのアクチン細胞骨格のダイナミクスを調べた。この実験系のアストロサイトは、SBF-SEMで観察した脳組織内のアストロサイトと同様に、細胞表面に重合アクチンが豊富な多数の細かい突起を持ち、この突起は伸長–退縮を繰り返しているが、移動する新生ニューロンが接触した部分では一次的にアクチン重合が抑制され、突起の形成が減少した。一方、Slit1を欠損する新生ニューロンが接しても、このような変化は見られなかった。よって、新生ニューロンはSlit1を用いてアストロサイトのアクチン重合を局所的に抑制して、形態変化を誘導していると考えられた。
Slit-Roboシグナルは、アクチン重合の主要な制御因子であるRhoファミリー低分子量G蛋白質RhoA・Rac1・Cdc42の活性調節により、細胞内のアクチン動態に影響を与える33)。これらの分子の活性を可視化するFRET(Fluorescence resonance energy transfer, 蛍光共鳴エネルギー移行)プローブ34)をアストロサイトに導入して新生ニューロンと共培養したところ、アストロサイト内のCdc42の活性が、新生ニューロンの接触によってSlit1依存的に低下することが分かった。アストロサイトに恒常活性化型Cdc42を導入すると、新生ニューロンの接触部でのアクチン重合低下は見られなくなり、新生ニューロンの移動速度も低下した。これらの結果から、新生ニューロンはSlit1を介してアストロサイト内のCdc42を不活性化しアクチン重合を低下させて、自身が移動しやすい細胞表面の形状を作り出していると考えられる。
梗塞巣に移動する新生ニューロンの数は、梗塞後2~3週をピークに減少していくが6, 29)、脳室下帯からの移動距離に着目した継時的変化については報告がなかった。そこで、脳梗塞後5週間までの新生ニューロンの分布の変化を調べたところ、多くの新生ニューロンは梗塞巣付近まで到達せず、アストロサイトが活性化した領域の外縁で移動を停止していた。また、新生ニューロンにおけるSlit1の発現レベルも、脳室下帯から遠ざかるとともに減少していた。そこで、新生ニューロンのSlit1発現を増加させることによって梗塞巣付近への新生ニューロンの移動を促進できるのではないかと考えて、レンチウィルスベクターを用いたSlit1強制発現実験を行なった。単離した脳室下帯細胞にSlit1発現ウィルスベクターを感染させたのち、梗塞後の脳内に移植したところ、新生ニューロンの移動速度が増加し、最終的に線条体外側の梗塞巣近くに定着する細胞の割合も増えた(図5)。これらの一部は、線条体の投射性ニューロンと同様の蛋白質の発現パターンや電気生理学的特徴を示し、線条体や、線条体ニューロンの主要な投射先のひとつである淡蒼球で、シナプスを形成していた。
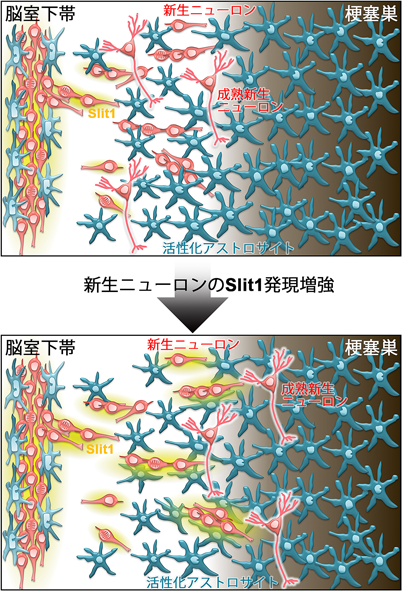
線条体から淡蒼球への軸索の投射には空間的な規則性が存在し、内側ニューロンは淡蒼球の内側に、外側ニューロンは淡蒼球外側に投射している35)。移植片由来ニューロンの軸索投射は、コントロール群では内側淡蒼球に偏在していたが、新生ニューロンの移動が促進されたSlit1強制発現群では淡蒼球外側まで広範囲にみられ、行動試験で定量評価した梗塞による神経機能障害も有意に改善した。この効果は脳梗塞2ヶ月後にも維持されていたが、遺伝学的手法36)を用いてニューロンに分化した移植細胞を除去すると消失したことから、Slit1を導入した移植細胞由来の新生ニューロンが神経機能改善を担っていたと考えられる。
最後に、神経機能と新生ニューロンの組織学的所見との関連性を検索した。細胞移植1ヶ月後の神経機能の改善度は、梗塞巣付近で定着した新生ニューロンの割合と比較的高い相関が見られたが、定着した新生ニューロンの総数とは関連が見られなかった。これらの結果から、新生ニューロンの移動促進により、梗塞巣付近に定着して成熟するニューロンが増加して、神経回路の再生が促進されたと考えられる。
上述した研究成果から、新生ニューロンの移動を促進して定着位置を変化させることで、神経機能の再生効率も向上することが示唆される。しかし、その神経回路レベルの詳細なメカニズムは未解明である。近年、脳梗塞の治療においても、細胞移植を中心とする再生医学的アプローチの研究が進められている。ドパミン分泌ニューロンなど一部の細胞を除けば、傷害脳内で神経回路に組み込まれて損傷した回路を補完することができなければ、機能的な再生を誘導することはできない。しかし、成体脳内で新生ニューロンの分化、生存、神経突起の発達がどのように制御され、既存の神経回路にどのような影響を与えるかはほとんど分かっていないのが現状である。出生後に発達する多種類のグリア細胞が関与するこの過程は、単なる発生の模倣とは考えがたく、医療応用の可能性のみならず、これから学術的な発展が期待される研究分野でもある。
神経科学分野の研究全般と同様に、脳梗塞後のニューロン再生の研究のほとんどは、マウスやラットを用いて行われている。しかし、患者の死後脳組織を用いた研究において、ヒトでも脳梗塞後に脳室下帯における新生ニューロンの産生が亢進し37, 38)、脳梗塞巣付近に新生ニューロンが出現することから39)、げっ歯類と同様にヒトの脳にも内在性のニューロン再生機構が潜在することが示唆されている。一方、最近の研究において、ヒトの脳室下帯のニューロン新生は新生児期以降に急速に減少して、生後1年半までにほとんど見られなくなることが明らかになった40, 41)。非侵襲的に新生ニューロンを標識し、その挙動を追跡する研究手法が存在しないため、病態下の成人の脳内で産生され、移動・成熟する新生ニューロンが存在するのかは現状では明らかになっていない。寿命も脳のサイズも大きく異なるヒトの脳内で新生ニューロンがどのようにふるまい、神経回路の可塑性や再生に関与するのかを明らかにすることは、基礎研究分野・臨床分野の研究者が連携して取り組むべき大きな課題であると考えている。
この度は、私の研究を日本神経化学会・優秀賞にご選出頂き、誠にありがとうございました。日本神経化学会は、私が精神科臨床医をしていた頃に基礎研究を開始して初めて入会した学会で、大会のたびに仲間と励ましあい、先生方にご指導頂くのを楽しみにしてきました。このような栄誉ある賞とともに、私の研究内容を本誌にご紹介する機会をくださった関係者の皆様に、心より御礼申し上げます。
臨床医時代の恩師である神庭重信先生(九州大学)、大学院生の私を研究室に加えてくださった岡野栄之先生(慶應義塾大学)、そして岡野研時代から現在まで15年にわたって辛抱強く、研究の楽しさ、厳しさを教えてくださった澤本和延先生(名古屋市立大学)に、この場を借りて深く感謝申し上げます。
1) Ihrie RA, Alvarez-Buylla A. Lake-front property: A unique germinal niche by the lateral ventricles of the adult brain. Neuron, 70(4), 674–686 (2011).
2) Lim DA, Alvarez-Buylla A. The adult Ventricular-Subventricular Zone (V-SVZ) and Olfactory Bulb (OB) neurogenesis. Cold Spring Harb Perspect Biol, 8(5), a018820 (2016).
3) Kaneko N, Sawada M, Sawamoto K. Mechanisms of neuronal migration in the adult brain. J Neurochem, 141(6), 835–847 (2017).
4) Arvidsson A, Collin T, Kirik D, Kokaia Z, Lindvall O. Neuronal replacement from endogenous precursors in the adult brain after stroke. Nat Med, 8(9), 963–970 (2002).
5) Parent JM, Vexler ZS, Gong C, Derugin N, Ferriero DM. Rat forebrain neurogenesis and striatal neuron replacement after focal stroke. Ann Neurol, 52(6), 802–813 (2002).
6) Yamashita T, Ninomiya M, Hernandez Acosta P, Garcia-Verdugo JM, Sunabori T, Sakaguchi M, Adachi K, Kojima T, Hirota Y, Kawase T, Araki N, Abe K, Okano H, Sawamoto K. Subventricular zone-derived neuroblasts migrate and differentiate into mature neurons in the post-stroke adult striatum. J Neurosci, 26(24), 6627–6636 (2006).
7) Lindvall O, Kokaia Z. Neurogenesis following Stroke Affecting the Adult Brain. Cold Spring Harb Perspect Biol, 7(11), a019034 (2015).
8) Cooper JA. Cell biology in neuroscience: Mechanisms of cell migration in the nervous system. J Cell Biol, 202(5), 725–734 (2013).
9) Marin O, Valiente M, Ge X, Tsai LH. Guiding neuronal cell migrations. Cold Spring Harb Perspect Biol, 2(2), a001834 (2010).
10) Nishimura YV, Nabeshima YI, Kawauchi T. Morphological and molecular basis of cytoplasmic dilation and swelling in cortical migrating neurons. Brain Sci, 7(7), (2017).
11) Lois C, Garcia-Verdugo JM, Alvarez-Buylla A. Chain migration of neuronal precursors. Science, 271(5251), 978–981 (1996).
12) Chazal G, Durbec P, Jankovski A, Rougon G, Cremer H. Consequences of neural cell adhesion molecule deficiency on cell migration in the rostral migratory stream of the mouse. J Neurosci, 20(4), 1446–1457 (2000).
13) Belvindrah R, Hankel S, Walker J, Patton BL, Muller U. Beta1 integrins control the formation of cell chains in the adult rostral migratory stream. J Neurosci, 27(10), 2704–2717 (2007).
14) Yagita Y, Sakurai T, Tanaka H, Kitagawa K, Colman DR, Shan W. N-cadherin mediates interaction between precursor cells in the subventricular zone and regulates further differentiation. J Neurosci Res, 87(15), 3331–3342 (2009).
15) Fujioka T, Kaneko N, Ajioka I, Nakaguchi K, Omata T, Ohba H, Fässler R, García-Verdugo JM, Sekiguchi K, Matsukawa N, Sawamoto K. beta1 integrin signaling promotes neuronal migration along vascular scaffolds in the post-stroke brain. EBioMedicine, 16, 195–203 (2017).
16) Kojima T, Hirota Y, Ema M, Takahashi S, Miyoshi I, Okano H, Sawamoto K. Subventricular zone-derived neural progenitor cells migrate along a blood vessel scaffold toward the post-stroke striatum. Stem Cells, 28(3), 545–554 (2010).
17) Snapyan M, Lemasson M, Brill MS, Blais M, Massouh M, Ninkovic J, Gravel C, Berthod F, Götz M, Barker PA, Parent A, Saghatelyan A. Vasculature guides migrating neuronal precursors in the adult mammalian forebrain via brain-derived neurotrophic factor signaling. J Neurosci, 29(13), 4172–4188 (2009).
18) Bolteus AJ, Bordey A. GABA release and uptake regulate neuronal precursor migration in the postnatal subventricular zone. J Neurosci, 24(35), 7623–7631 (2004).
19) Mason HA, Ito S, Corfas G. Extracellular signals that regulate the tangential migration of olfactory bulb neuronal precursors: Inducers, inhibitors, and repellents. J Neurosci, 21(19), 7654–7663 (2001).
20) Kaneko N, Marin O, Koike M, Hirota Y, Uchiyama Y, Wu JY, Lu Q, Tessier-Lavigne M, Alvarez-Buylla A, Okano H, Rubenstein JL, Sawamoto K. New neurons clear the path of astrocytic processes for their rapid migration in the adult brain. Neuron, 67(2), 213–223 (2010).
21) Nguyen-Ba-Charvet KT, Picard-Riera N, Tessier-Lavigne M, Baron-Van Evercooren A, Sotelo C, Chedotal A. Multiple roles for slits in the control of cell migration in the rostral migratory stream. J Neurosci, 24(6), 1497–1506 (2004).
22) Ohab JJ, Fleming S, Blesch A, Carmichael ST. A neurovascular niche for neurogenesis after stroke. J Neurosci, 26(50), 13007–13016 (2006).
23) Robin AM, Zhang ZG, Wang L, Zhang RL, Katakowski M, Zhang L, Wang Y, Zhang C, Chopp M. Stromal cell-derived factor 1alpha mediates neural progenitor cell motility after focal cerebral ischemia. J Cereb Blood Flow Metab (Nihongoban), 26(1), 125–134 (2006).
24) Yan YP, Lang BT, Vemuganti R, Dempsey RJ. Osteopontin is a mediator of the lateral migration of neuroblasts from the subventricular zone after focal cerebral ischemia. Neurochem Int, 55(8), 826–832 (2009).
25) Yan YP, Sailor KA, Lang BT, Park SW, Vemuganti R, Dempsey RJ. Monocyte chemoattractant protein-1 plays a critical role in neuroblast migration after focal cerebral ischemia. J Cereb Blood Flow Metab (Nihongoban), 27(6), 1213–1224 (2007).
26) Belayev L, Alonso OF, Busto R, Zhao W, Ginsberg MD. Middle cerebral artery occlusion in the rat by intraluminal suture. Neurological and pathological evaluation of an improved model. Stroke, 27(9), 1616–1622, discussion, 23 (1996).
27) Longa EZ, Weinstein PR, Carlson S, Cummins R. Reversible middle cerebral artery occlusion without craniectomy in rats. Stroke, 20(1), 84–91 (1989).
28) Sofroniew MV. Astrogliosis. Cold Spring Harb Perspect Biol, 7(2), a020420 (2015).
29) Thored P, Arvidsson A, Cacci E, Ahlenius H, Kallur T, Darsalia V, Ekdahl CT, Kokaia Z, Lindvall O. Persistent production of neurons from adult brain stem cells during recovery after stroke. Stem Cells, 24(3), 739–747 (2006).
30) Kaneko N, Herranz-Perez V, Otsuka T, Sano H, Ohno N, Omata T, Nguyen HB, Thai TQ, Nambu A, Kawaguchi Y, García-Verdugo JM, Sawamoto K. New neurons use Slit-Robo signaling to migrate through the glial meshwork and approach a lesion for functional regeneration. Sci Adv, 4(12), eaav0618 (2018).
31) Riedl J, Crevenna AH, Kessenbrock K, Yu JH, Neukirchen D, Bista M, Bradke F, Jenne D, Holak TA, Werb Z, Sixt M, Wedlich-Soldner R. Lifeact: A versatile marker to visualize F-actin. Nat Methods, 5(7), 605–607 (2008).
32) Yamashiro S, Mizuno H, Smith MB, Ryan GL, Kiuchi T, Vavylonis D, Watanabe N. New single-molecule speckle microscopy reveals modification of the retrograde actin flow by focal adhesions at nanometer scales. Mol Biol Cell, 25(7), 1010–1024 (2014).
33) Wong K, Ren XR, Huang YZ, Xie Y, Liu G, Saito H, Tang H, Wen L, Brady-Kalnay SM, Mei L, Wu JY, Xiong WC, Rao Y. Signal transduction in neuronal migration: Roles of GTPase activating proteins and the small GTPase Cdc42 in the Slit-Robo pathway. Cell, 107(2), 209–221 (2001).
34) Yoshizaki H, Ohba Y, Kurokawa K, Itoh RE, Nakamura T, Mochizuki N, Nagashima K, Matsuda M. Activity of Rho-family GTPases during cell division as visualized with FRET-based probes. J Cell Biol, 162(2), 223–232 (2003).
35) Voorn P, Vanderschuren LJ, Groenewegen HJ, Robbins TW, Pennartz CM. Putting a spin on the dorsal-ventral divide of the striatum. Trends Neurosci, 27(8), 468–474 (2004).
36) Kobayakawa K, Kobayakawa R, Matsumoto H, Oka Y, Imai T, Ikawa M, Okabe M, Ikeda T, Itohara S, Kikusui T, Mori K, Sakano H. Innate versus learned odour processing in the mouse olfactory bulb. Nature, 450(7169), 503–508 (2007).
37) Marti-Fabregas J, Romaguera-Ros M, Gomez-Pinedo U, Martinez-Ramirez S, Jimenez-Xarrie E, Marin R, Martí-Vilalta JL, García-Verdugo JM. Proliferation in the human ipsilateral subventricular zone after ischemic stroke. Neurology, 74(5), 357–365 (2010).
38) Minger SL, Ekonomou A, Carta EM, Chinoy A, Perry RH, Ballard CG. Endogenous neurogenesis in the human brain following cerebral infarction. Regen Med, 2(1), 69–74 (2007).
39) Jin K, Wang X, Xie L, Mao XO, Zhu W, Wang Y, Shen J, Mao Y, Banwait S, Greenberg DA. Evidence for stroke-induced neurogenesis in the human brain. Proc Natl Acad Sci USA, 103(35), 13198–13202 (2006).
40) Paredes MF, James D, Gil-Perotin S, Kim H, Cotter JA, Ng C, Sandoval K, Rowitch DH, Xu D, McQuillen PS, Garcia-Verdugo JM, Huang EJ, Alvarez-Buylla A. Extensive migration of young neurons into the infant human frontal lobe. Science, 354(6308), aaf7073 (2016).
41) Sanai N, Nguyen T, Ihrie RA, Mirzadeh Z, Tsai HH, Wong M, Gupta N, Berger MS, Huang E, Garcia-Verdugo JM, Rowitch DH, Alvarez-Buylla A. Corridors of migrating neurons in the human brain and their decline during infancy. Nature, 478(7369), 382–386 (2011).

This page was created on 2019-11-13T16:59:47.155+09:00
This page was last modified on 2019-12-16T11:48:23.000+09:00
このサイトは(株)国際文献社によって運用されています。