脳梗塞後慢性期における制御性T細胞による組織修復メカニズムの解明
慶應義塾大学医学部微生物学免疫学教室
発行日:2019年12月30日Published: December 30, 2019
© 2019 日本神経化学会© 2019 The Japanese Society for Neurochemistry
脳血管障害(脳卒中)は日本人の死因の第3位で年間11万人弱が死亡し、患者総数は100万人を超えている。脳卒中のなかでも、脳の血管が詰まるなど血流が減少することによって、酸素や栄養が不足して脳組織が壊死に至る脳梗塞が約75%を占めている。脳梗塞は死亡率が高いだけでなく後遺症が重く、患者の生活の質(QOL)が非常に低下する。発症後数時間以内の脳梗塞であれば、詰まった血管の再開通(血栓溶解または血栓除去)によって神経症状や後遺症の改善が期待できるが、そのような早期に発見できるケースは限られており、それ以降の有効な治療法に乏しい。
これまでに、著者らの研究室では、マクロファージを中心とした脳梗塞発症後の炎症プロセスを明らかにしてきた。発症1日目には炎症性のマクロファージが梗塞部位に浸潤し、死細胞由来の物質を認識して炎症性サイトカインを放出する1, 2)。その後3日目にγδT細胞が浸潤しIL-17を放出して神経細胞死が亢進する3)。それ以降はマクロファージが修復性に転換し炎症物質を除去、およそ1週間で炎症反応が収束する4)。しかし、それ以降の慢性期において炎症は収まっていると考えられてきたため、免疫細胞の役割についてはほとんど解析されていなかった。本稿では特に神経症状の緩和に重要な脳制御性T細胞について概説する5)。
制御性T細胞(Treg)は、転写因子Forkhead box P3(Foxp3)を主要なマスター転写因子とし、自己抗原、共生細菌由来抗原、および外来抗原など多様な抗原に対して過剰な免疫応答を抑制することで、生体内の恒常性の維持に働いている6)。これまでの研究ではTregが抑制する相手としては主に活性化されたエフェクターT細胞や樹状細胞、マクロファージなどの免疫細胞が中心であった。しかし近年、定常状態および傷害時に非リンパ組織に局在し、非免疫細胞に作用するTreg細胞が注目を集めている。それらは組織Tregと呼ばれ共通の性質を示すと同時に、それぞれの組織に特化した特徴を有する(図1)7)。これまで脳内のTregは脳や脊髄の損傷後の炎症や多発性硬化症(MS)のマウスモデルである実験的自己免疫性脳脊髄炎(EAE)で解析されて来たが、組織修復に関係する脳内Tregの存在は不明であった。
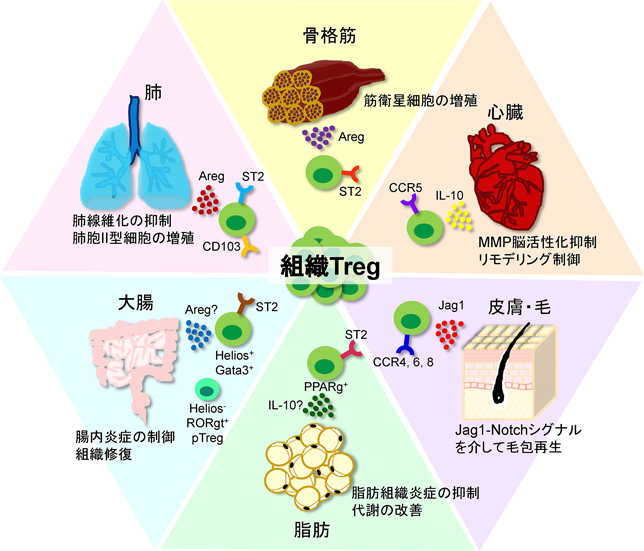
様々な組織で組織特異的な特徴を有する組織Tregが存在する。
実験的脳虚血再灌流(MCAO)モデル(脳梗塞モデル)では、発症2週目以降の慢性期の脳内に制御性T細胞が集積することが見出された(図2)。これまでに脳梗塞急性期(1週間以内)にTregが少数脳内に認められ、脳梗塞後の神経障害を抑制するとされてきたが、浸潤する細胞数は極めて少なく、組織Tregとしての性質を示さずにIL-10を産生するなどバイスタンダーとして若干の寄与をするのみである。しかし慢性期のTregはその数10倍以上存在し、脳梗塞巣の内部のみならず梗塞部位周辺に局在し、アストロサイトや生き残った神経細胞と近接していることが示された。DEREGマウス(ジフテリア毒素DTによってTregを一過性に除去できるマウス)を用いてTregを除去すると神経症状が悪化した。逆にRag欠損マウスやCD3ε欠損マウスのようなT細胞が存在しないマウスにTregを戻すと神経症状が改善された。これらのことから脳梗塞慢性期には脳内にTregが大量に浸潤し神経症状の改善に重要な役割を果たしていることが示された。

脳梗塞慢性期には梗塞巣とその周囲にT細胞が集積している。CD4+T細胞のうち約半数がTregであり、脳Tregは発症後2週間程度で脳内に浸潤し増加した。
そこで次に脳Tregの増幅機構を調べた。IL-2はすべてのTregの生存、増殖で必須なサイトカインとして知られているが、抗IL-2抗体の投与によって脳Tregの数は減少した。また、脳TregはIL-33受容体(ST2)を高発現することからIL-33の関与が示唆された。そこでIL-33欠損マウス、ST2欠損マウスを調べたところ脳Tregの数は減少した。よって脳Tregの増幅にはIL-2、IL-33が必須といえる。ちなみにIL-33の発現は脳梗塞によって増加し、オリゴデンドロサイトとアストロサイトが主な産生源である。IL-33を欠損するマウスは、修復性のM2マクロファージ関連遺伝子の誘導が減少し、神経損傷後の回復ができなくなることが報告されており、これらのIL-33の修復機能に脳Tregが関与する可能性は高い。
次になぜ脳Tregは脳指向性なのかを調べた。通常リンパ球の組織への遊走にはケモカインとその受容体が関与する。Tregはケモカイン受容体のうちCCR6とCCR8を高発現し、脳梗塞巣で上昇するCCL20やCCL1依存的に脳内に浸潤した。逆にCCL20やCCL1を脳室投与するとTregが増加し、神経症状も改善された。
遺伝子発現解析からも脳Tregは他の組織Tregに類似していることが示された。しかし脳Tergにはそれらにない特徴的な遺伝子発現も見られた。特に通常Tリンパ球に見られない神経関連遺伝子がいくつか発現している。脳TregはcAMPを上昇させるセロトニン受容体7(Htr7)を発現していた(図3a)。cAMPはTregの増殖を促進すると同時に機能を増強させることが知られている。予想通り単離された脳TregはHtr7依存的に増殖・活性化された。脳梗塞慢性期にセロトニンや選択的セロトニン再取り込み阻害薬(SSRI)を投与すると脳Tregが増加し神経症状が改善した(図3b)。よって脳Tregは脳という特殊な環境に順応した機能を獲得しており、神経伝達物質によって刺激を受ける増幅するという非常に興味深い性質を有している。どのような機構で脳Tregは神経関連遺伝子を発現するのかは解明すべき今後の重要な課題である。
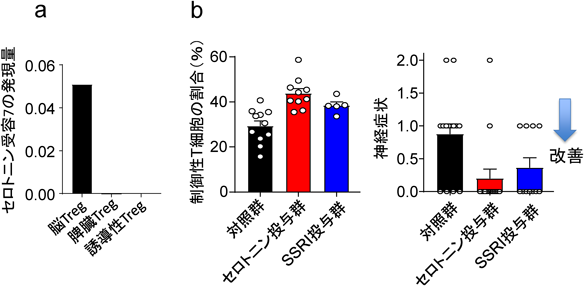
(a)脳Tregはセロトニン受容体7を発現しているがそれ以外の組織のTregでは発現していない。(b)セロトニンやSSRI(選択的セロトニン再取り込み阻害薬:Fluoxetine)を脳梗塞慢性期(発症7日後から)投与すると、脳Tregは増殖し、神経症状も改善した。
ではどのような仕組みで脳Tregは脳機能改善に寄与しているのであろうか?脳内Tregを除去、あるいは減少させるとアストロサイトの過剰な活性化が認められ、運動野の神経細胞がアポトーシスを起こしていた。これがTreg除去による運動機能回復の遅れに繋がっていると考えられる。アストロサイトの活性化は瘢痕形成を誘導し神経細胞を炎症から隔離するために必要であるが、過剰に活性化されたアストロサイトは神経毒性因子を産生し、むしろ神経細胞を傷害したり、神経伸長を阻害したりする8)。脳Tregはアストロサイトの過剰な活性化(アストログリオーシス)を抑制していることがわかった。アストロサイトの活性化にはIL-6などの炎症性サイトカインが重要である。試験管内で脳Tregと活性化したミクログリアやアストロサイトを共培養すると、IL-6の産生が抑制されることがわかった。またTregの移入によりアストロサイトの過剰な活性化が抑制され、IL-6の下流で活性化されるSTAT3のリン酸化が抑制され、神経症状が改善した。脳Tregの特徴のひとつはAregを強く産生することであるが、AregはマクロファージにおいてIL-6やTNF-αなどの炎症性サイトカインの産生を抑制することが知られている。Aregの脳室内投与により、アストロサイトの活性化が抑制され、神経症状が改善した。試験管内の実験で抗Areg抗体を投与すると脳TregによるミクログリアやアストロサイトからのIL-6産生抑制が部分的に解除された。さらに個体でもAregを欠損するTregでは野生型Tregで認められたアストロサイトの抑制や神経症状の改善が認められなかった。AregがIL-6産生を阻害する分子機構は不明であるものの、Aregは脳Tregの重要な機能分子と言える。またAregは神経幹細胞の増殖に直接的に働いている可能性もある9)。
脳梗塞後の脳Tregの意義や特徴、脳Tregによる組織修復機構ついて概説した(図4)。組織損傷後の炎症の慢性期にTregが修復に働いているのは脳だけではない。多くの組織で一見炎症が治まったように見えてもTregが組織修復や恒常性の維持に働いている可能性がある。組織におけるTregの生存や増殖をもたらす自己抗原やTregを刺激するリガンドは、組織特異的なTregの機能に重要である可能性が高い。抗原やリガンドの探索も含め、Tregの組織特異的なフェノタイプを解析することによって、組織特異的なTregを誘導、増殖することができれば、Tregを用いた組織特異的な治療法の開発につながることが期待される。
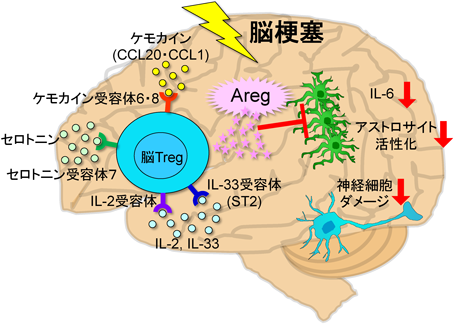
脳Tregは脳梗塞慢性期にケモカイン依存的に脳内に浸潤する。インターロキン(IL-2, IL-33)やセロトニンによって増殖し、アンフィレグリン(Areg)を産生する。Aregはインターロイキン6(IL-6)の産生を抑制することでアストロサイトの過剰な活性化を抑え、神経細胞を守る。
本研究を遂行するにあたり、多大なるご指導を賜りました慶應義塾大学の吉村昭彦教授および研究室の皆様、東京都医学総合研究所の七田崇プロジェクトリーダーに心より感謝申し上げます。また、ご助言を賜りました慶應義塾大学の田中謙二先生、名古屋市立大学の澤本和延先生に厚く御礼申し上げます。最後に、本稿執筆の機会を与えてくださいました日本神経化学会奨励賞選考委員の先生方、関係者の先生方に厚く御礼申し上げます。
1) Shichita T, Hasegawa E, Kimura A, Morita R, Sakaguchi R, Takada I, Sekiya T, Ooboshi H, Kitazono T, Yanagawa T, Ishii T, Takahashi H, Mori S, Nishibori M, Kuroda K, Akira S, Miyake K, Yoshimura A. Peroxiredoxin family proteins are key initiators of post-ischemic inflammation in the brain. Nat Med, 18(6), 911–917 (2012).
2) Ito M, Shichita T, Okada M, Komine R, Noguchi Y, Yoshimura A, Morita R. Bruton’s tyrosine kinase is essential for NLRP3 inflammasome activation and contributes to ischaemic brain injury. Nat Commun, 6(1), 7360 (2015).
3) Shichita T, Sugiyama Y, Ooboshi H, Sugimori H, Nakagawa R, Takada I, Iwaki T, Okada Y, Iida M, Cua DJ, Iwakura Y, Yoshimura A. Pivotal role of cerebral interleukin-17-producing gammadeltaT cells in the delayed phase of ischemic brain injury. Nat Med, 15(8), 946–950 (2009).
4) Shichita T, Ito M, Morita R, Komai K, Noguchi Y, Ooboshi H, Koshida R, Takahashi S, Kodama T, Yoshimura A. MAFB prevents excess inflammation after ischemic stroke by accelerating clearance of damage signals through MSR1. Nat Med, 23(6), 723–732 (2017).
5) Ito M, Komai K, Mise-Omata S, Iizuka-Koga M, Noguchi Y, Kondo T, Sakai R, Matsuo K, Nakayama T, Yoshie O, Nakatsukasa H, Chikuma S, Shichita T, Yoshimura A. Brain regulatory T cells suppress astrogliosis and potentiate neurological recovery. Nature, 565(7738), 246–250 (2019).
6) Ito M, Iizuka-Koga M, Ando M, Yoshimura A. Development and functional modulation of regulatory T cells by transcription factors and epigenetics. Cornea, 37(Suppl 1), S42–S49 (2018).
7) Panduro M, Benoist C, Mathis D. Tissue tregs. Annu Rev Immunol, 34(1), 609–633 (2016).
8) Liddelow SA, Guttenplan KA, Clarke LE, Bennett FC, Bohlen CJ, Schirmer L, Bennett ML, Munch AE, Chung WS, Peterson TC, Wilton DK, Frouin A, Napier BA, Panicker N, Kumar M, Buckwalter MS, Rowitch DH, Dawson VL, Dawson TM, Stevens B, Barres BA. Neurotoxic reactive astrocytes are induced by activated microglia. Nature, 541(7638), 481–487 (2017).
9) Kimura H, Schubert D. Schwannoma-derived growth factor promotes the neuronal differentiation and survival of PC12 cells. J Cell Biol, 116(3), 777–783 (1992).

This page was created on 2019-11-11T19:27:52.107+09:00
This page was last modified on 2019-12-16T11:51:42.000+09:00
このサイトは(株)国際文献社によって運用されています。