神経化学的方法に基づいた合理的設計による新規超高速多色カルシウムセンサーの開発
Department of Bioengineering, Stanford University School of Medicine
発行日:2019年12月30日Published: December 30, 2019
© 2019 日本神経化学会© 2019 The Japanese Society for Neurochemistry
脳は複数の異なる神経細胞種がそれぞれ時空間的に決まった順序で発火し、正常な認知・学習機能を発揮すると考えられている。特に、脳情報は神経ネットワークにおける特定の細胞種の神経発火パターン(周波数、発火タイミング、および異なる細胞同士の同期発火)にコードされていると考えられているが、個別の行動と神経発火パターンの関係は不明である。その主要な理由として計測手法が未発達であることが挙げられる。これまでの神経活動(神経発火、シナプス活動)計測には、電気生理学的手法により神経発火を計測する手法が汎用されてきた。この手法は高い時間分解能を有しているものの、細胞種および細胞の空間情報を厳密に同定することはできず、活動をしていない細胞に至ってはその存在を検出できない。近年、この脳情報を読み解くのに、発火に伴い細胞外のカルシウム(Ca2+)が細胞内に流入することを利用して、蛍光Ca2+センサーを用いた神経活動イメージング法が急速に普及しつつある。これを応用して、空間情報を保持したまま、数万個もの大規模な神経活動を単一細胞レベルで同時に記録することが可能である。その中でも、特定の神経細胞種に発現させることができ、長期観察可能であるため、GCaMPをはじめとする遺伝子にコードされたCa2+センサー(Genetically Encoded Calcium Indicator、以下GECIと略)が広く用いられている。しかしながら、従来の生体内で実用的なGECIは緑色のみであるため、複数の神経細胞種の発火を同時計測するには困難であった。加えて、従来のGECIは入力の有無のみを検出しており、個々の神経発火パターン(発火頻度、回数)を読み取るには不十分であった。
本稿では脳情報を精密に解読するための新規超高速多色Ca2+センサーの開発とその応用例について過去の報告と筆者らの研究成果を紹介する。
現在では神経科学研究に欠かせない技術となっているCa2+イメージングによる神経活動計測は1982年にRoger Tsienらが低分子蛍光色素1)を開発したことに端を発し、1997年に同研究室の宮脇博士らが2つの蛍光タンパク質のスペクトルの変化を利用したFRET型のcameleon2)を開発したことによりGECIの歴史が始まった。この知見を基に、2001年に中井博士らが緑色単色のG-CaMP3)を開発したことにより、標準的な顕微鏡システムにおいてCa2+濃度変化の計測を可能にした。GCaMPは改変GFPとカルモジュリン(CaM)およびCa2+/CaM結合領域であるMLCK由来のM13配列からなり、Ca2+濃度の上昇に伴い蛍光強度が増加する。さらに、中井グループ、Janeliaがこれを改良し、GCaMP3–8およびjGCaMP7と順次開発し4–9)、Ca2+フリーとCa2+飽和状態のダイナミックレンジを最大化させる努力してきた。これにより、哺乳類in vivoで発火の有無を検出できるようになったが、未だ発火パターンを精密に解読するのが困難であった。
近年開発されたGCaMPはCa2+飽和状態の蛍光強度が元となるEGFPと同程度の明るさまでに至り、(改変EGFPを使用している限り)上限の蛍光強度を改善する余地はほぼないと考えられる。また、Ca2+フリー状態の蛍光強度においても発火が無い静的な状態において蛍光強度を検出できないとΔF/Fを計算できないため(この場合、ΔF/Fは無限値をとる)、下限値の改善にも限界がある。では、どの点に改善の余地があるだろうか。①In vivoにおける細胞内Ca2+濃度は数十nMと考えられているので、単一発火に伴う微弱なCa2+濃度変化を捉えるにはこの濃度に対応した高感度であることが必要がある(図1a)。②また、単一発火を検出し、かつ20発のような連続発火を違いとして検出するような発火回数と蛍光変化が線形に変化する必要がある。これを実現するために、Ca2+濃度と蛍光強度の変化の関係を示すHill係数が『1』のGECIを作出することが必要である(図1b)。しかしながら、従来のGECIはHill係数が2–3付近であることから、複数の発火に対し非線形的に蛍光強度が変化し、かつ、広範囲のCa2+濃度を検出するのが困難であった。
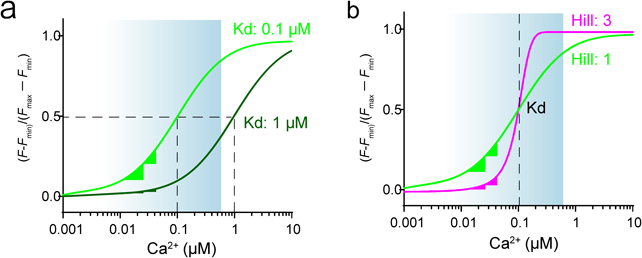
(a)見たい現象のCa2+濃度変化の範囲に合わせて、適切な感度のCa2+センサーを選択する必要がある。神経発火に伴うCa2+濃度変化なら、Kd: 100–200 nMの範囲が至適レンジと考えられる。Kd値が100 nM(緑)と1 µMで静的なCa2+濃度状態から発火に伴うCa2+濃度変化の蛍光変化率が全く異なる。青色のグラデーションは予想される神経発火に伴うCa2+濃度変化の範囲。 (b)同じKd値であっても、Hill係数の違いにより、検出できるCa2+濃度変化の範囲が異なる。Hill係数が1に近いほど、より広範囲かつ線形な変化になる。Hill係数が高くなるほど、より狭い範囲のCa2+変化を検出し、Hill係数が高いと、2つの異なる静的なCa2+濃度状態から絶対値のCa2+濃度変化が同じでも、全く異なる蛍光強度変化を引き起こす(マゼンタ)。一方、Hill係数が1だと、同じ蛍光変化率である(緑)。青色のグラデーションは予想される神経発火に伴うCa2+濃度変化の範囲。
また、従来の最速のGECIでも単一発火に伴うCa2+立ち上がりが60ミリ秒、Ca2+減衰は200ミリ秒程度と遅いので、高頻度の発火を解読するためには蛍光変化の速度を早くする必要がある。低分子蛍光色素を含めたCa2+センサー全体においても、従来のセンサーはパルアルブミン(PV)抑制性細胞のような100 Hzで高頻度発火する細胞の活動を読み取ることが課題として残されていた。
一方、緑色GECIの有用性が認知されるに伴い、Campbell・永井グループが2011年に初めて赤色GECI(R-GECO)10)の開発に成功し、中井グループおよびJanaliaがこれを改良し、R-CaMP1.0711)、RCaMP112)を開発した。しかしながら、これらの赤色GECIは脳スライス標本においてまでしか発火を検出できていなかったことから、生体内において複数細胞種の神経発火を同時にかつ精密に解読することが困難であった。これらの課題を解決するために①多色化、②Ca2+に対する蛍光変化が早く、③Ca2+高親和性かつ、④発火回数と蛍光変化に線形の関係があるGECIを開発することが望ましいと考えた。
上述したように、GECIの性能を決めるものとして4条件(ダイナミックレンジ、高感度、Hill係数、キネティクス)がある。これまでのアミノ酸変異をランダムに導入するスクリーニングでは全条件が変動するので、これを兼ね備えるセンサーを見出すのは困難である7, 10)。著者らは4条件の内1つでも上記条件を制御可能にすれば、従来のスクリーニング法より効率的に有用センサーを見出すことができると考えた。筆者らはこれまでに生化学的手法によりCaMKKはCa2+とカルモジュリン(CaM)との複合体に対し高親和性かつHill係数が低いことを示唆する結果を見出したことから、上記2条件はあらかじめ備わっていることが示唆される。これを検証するため、従来の最高性能の赤色GECIのR-CaMP1.07を骨格として用い、M13配列をckkap配列と置換した。さらに、高いダイナミックレンジを最適化させるために、合計で150個の候補となる変異体をin vitroにおいてスクリーニングをした。その結果、既存の赤色Ca2+センサーに比べ感度が3倍も向上し、かつ、Hill係数が1付近であることから線形性が圧倒的に高い赤色Ca2+センサーを作出し、R-CaMP2と命名した13)。予想外のボーナスとして、皮質スライス標本において、従来の高速赤色GECI(R-CaMP1.07)の4倍の40 Hzもの高頻度の発火に追従する高速GECIの作出に成功した。また、In vivoにおいて、2光子イメージング法と電気生理法を同時に行い、R-CaMP2は入力回数と蛍光変化に強い線形の関係にあることを見出し(図2)、単一活動電位を十分に検出できる唯一の赤色GECIであることを示した。R-CaMP2は生体内で神経発火およびシナプス活動の計測を初めて可能とする赤色GECIであり、緑色GECIと組み合わせることで、マウス大脳皮質における興奮性錐体細胞(Pyr)と抑制性ソマトスタチン(SST)細胞の2つの異なる神経細胞種の神経活動を同時に計測することを可能にした。その結果、SST細胞同士の同期発火がSST-PyrおよびPyr細胞同士よりはるかに高いことを示し、従来の電気生理学的知見と一致することを確認した。この結果は2色のCa2+センサーを用いることにより、初めて2つの神経細胞種の関係を明らかにしたことになる。さらに、自由行動中の線虫の抑制性運動神経細胞にチャネルロドプシンと緑色GECIを発現させ、一方その投射先である筋細胞にR-CaMP2を発現させて、それぞれの活動を同時計測した。光照射により、抑制性運動神経では活動が上昇する一方、標的である筋細胞ではCa2+濃度の急速低下に伴い弛緩が生じ、それまで動いていた線虫を運動停止させることに成功した。このように、光遺伝学的行動制御を実施するときに、関連回路内における高い時間分解能での神経活動計測を実践することが可能となった。
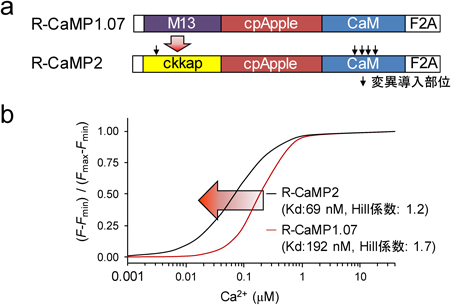
(a)従来の赤色Ca2+センサーR-CaMP1.07のCa2+/CaM結合配列(M13)を、R-CaMP2ではCAMKKのCa2+/CaM結合配列(ckkap)と置換した。矢印は変異を導入した部位。 (b) R-CaMP1.07およびR-CaMP2の滴定曲線。Ca2+に対し高親和性かつHill係数が1付近の赤色Ca2+センサーを作出することに成功した。文献13より改変・転載。
R-CaMP2開発の知見により、ckkap配列はGECIの性能を決める4条件(高感度、ダイナミックレンジ、キネティクス、Hill係数)のうち、3条件(高感度、キネティクス、Hill係数)制御可能であることが示唆された。それではckkap配列はどれくらい普遍性があるのか、他の色に適応して検討することにした。まず、既存の緑色GECIの中では最もHill係数が低いG-CaMP4.1(Hill係数=2.1)を骨格として、M13配列をckkap配列に置換した。さらに、リンカー配列に合計100個の変異を導入し、in vitroにてスクリーニングした。その結果、どの候補センサーも20–200 nMとCa2+高感度であり、Hill係数も1付近と線形性を保っていることから、我々の仮説を裏付ける結果となった(図3)。さらに、脳スライス標本にてスクリーニングした結果、これまでのCa2+センサーで最速の緑色GECIであるXCaMP-Gfおよび単一の活動電位に伴うCa2+応答が最大のダイナミックレンジを持つXCaMP-Gを開発した。これを基にし、構造情報を用いて多色化をするための変異を加えて、Hill係数がおよそ1の『XCaMP(それぞれXCaMP-B、XCaMP-Y、XCaMP-R)』を作成した14)。生体マウス脳におけるXCaMPの性能を検証するために、各XCaMPおよび従来のGECIを大脳皮質バレル野第2/3層錐体細胞にそれぞれ強制発現させ、2光子イメージング法と電気生理法を同時に行った。その結果、全てのXCaMPセンサーが神経発火回数と蛍光強度の変化の間に強い線形の関係を有することを見出し、単一の活動電位を鋭敏に検出することを示した。単一活動電位に対しXCaMP-Gf, XCaMP-Gは従来の最も高性能なGCaMP6fに比べ3–5倍も大きいダイナミックレンジを示し、立ち上がり、減衰キネティクスもそれぞれ4倍、1.2–1.5倍早くなった。この結果は、XCaMP-Gfが遺伝子にコードされた全てのCa2+センサーの中で世界最高速であることを意味する。XCaMP-YとXCaMP-RもXCaMP-Gfと同等な最高性能なダイナミックレンジおよびキネティクスを示した。
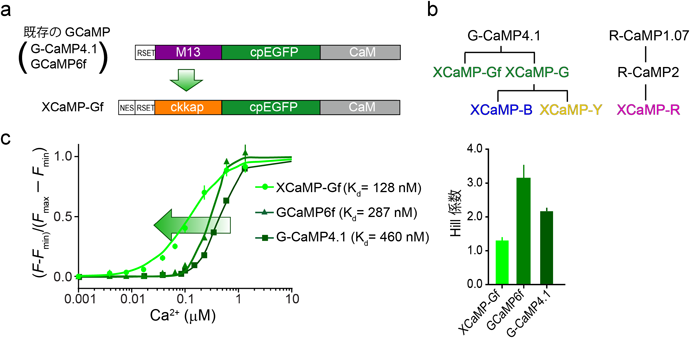
(a)従来の緑色GECI(G-CaMP4.1)のCa2+/CaM結合配列(M13)をCaMKKのCa2+/CaM結合配列(ckkap)に置換し、さらなる変異を追加することにより、Ca2+に対して高親和性の緑色GECIの作出に成功した。(b)XCaMPセンサーの系譜。(c)従来のGECIはCa2+濃度と蛍光変化の関係を示すHill係数が2付近またはそれ以上であったことから、複数の神経発火に対し非線形的に蛍光変化するのに対し、XCaMP-GfはHill係数を『1』に近づけたため、非常に高い線形性を示す。文献14より改変・転載。
次に、世界最速のXCaMP-Gfを用いることにより、マウス生体内において高頻度発火するパルアルブミン(PV)陽性抑制性神経細胞の発火パターンを読み取れるかを検討した。PV細胞にXCaMP-Gfまたは比較のためにGCaMP6fを発現させ、2光子イメージング法と電気生理法を同時に行った。Ca2+イメージングのデータから発火頻度を予測し、実際の電気生理のデータと答え合わせをすることにより、両者のGECIの性能を比較した。その結果、高頻度発火帯においてXCaMP-GfはGCaMP6fよりも発火頻度を有意に精度良く読み取れることを示された(図4a)。
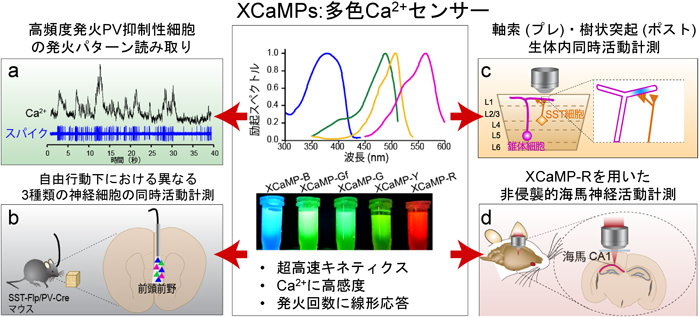
超高速・高感度・線形性を特徴とする多色Ca2+センサーの『XCaMP』センサーを開発したことにより、(a)高頻度発火PV抑制性細胞の発火パターンの解読、(b)マウス自由行動下における異なる3種類の神経細胞の同時計測、(c)軸索から樹状突起への神経情報が伝達する様子の同時活動計測、(d)非侵襲的に脳深部である海馬神経活動計測に成功した。文献14より改変・転載。
また、生体内で計測可能な初めての青色のXCaMP-Bを作成したことにより、自由行動下においてXCaMP-GfとXCaMP-Rを組み合わせて、異なる3種類の神経細胞種(興奮性錐体細胞、抑制性細胞のPVおよびSST細胞)の同時活動計測を試みた。自由行動下においてミリ秒単位の高速神経活動計測を可能とするファイバーフォトメトリー法を用いてマウス前頭前野における物体探索行動時の神経活動計測に初めて成功した(図4b)。その結果、抑制性PV細胞がSST細胞に300ミリ秒程先行して活動し、その相前後に興奮性錐体細胞が活動するという各細胞種が時間的に精緻に制御されていることを初めて示した。
さらに、従来測定可能な色域が限られていたため、樹状突起または軸索の一方のみしか活動計測できなかったので、両者の関係については不明であった。生体内で計測可能な初めての黄色のXCaMP-Yを作成したことにより、XCaMP-Rと組み合わせれば、一つの2光子励起波長でプレ・ポストシナプスといった小さい構造物を同時計測可能であることが期待される。興奮性細胞の樹状突起にXCaMP-Rを、抑制性SST細胞の軸索にXCaMP-Yを発現させ、時空間的な軸索・樹状突起間の発火制御測定を試みた(図4c)。その結果、SST抑制性軸索が活動した時のみ、空間的に隣接する興奮性樹状突起での活動が有意に低下した。時間的には、軸索が活性化する1秒前後で樹状突起の活動が有意に低下した。これは、抑制性シナプス伝達によるtranssynapticな時空間動態制御を可視化した最初の例となる。
最後に、XCaMP-Rが赤色と長波長シフトしているため、光の組織内散乱が少ないことを利用し、従来型の2光子顕微鏡を用いて従来の可視化対象上部の大脳皮質を一切除去せず、非侵襲的に海馬CA1領域の錐体細胞神経活動の直接計測を実現した(図4d)。感覚入力に伴った持続的な活動様式を観察することができました。以上のように、XCaMPを用いることにより、従来計測困難であった複数細胞種間の脳神経ダイナミクスを高時空分解能で明らかにする新規の方法論を提示したと言える。
脳の中でどのように発火パターンで情報処理をしているのかを解明するために、生きた個体内で神経活動を可視化するツールおよび顕微鏡技術の進歩はめざましい。2008年にノーベル化学賞を受賞したRoger Tsien博士らが低分子蛍光色素およびGFPについて成し遂げた偉業に端を発し、宮脇・Campbell・永井・中井らが継承してきたチャレンジに新たな展開をもたらすと考えられる。本研究では新規配列を用いて合理的に設計することにより、ダイナミックレンジを最大化するだけでなく、線形化・高速化Ca2+センサーを開発し、神経活動の有無だけでなく神経発火パターンを精密に計測することを可能にした。同時に、in vivoで計測可能な多色Ca2+センサーを開発し、従来では不可能であった2つ以上の異なる神経ネットワークの関係を解明することを可能にし、新たなツールキットを構築したと考えられる。この方法を用いることにより、神経発火パターンの解読による脳の作動原理をはじめとして、神経発火パターンの異常に起因する精神疾患の解明に役立つことが期待される。行動中の神経発火パターンを計測できるようになれば、行動と神経発火パターンの因果関係を求めたくなる。様々なスペクトルを持つオプシンと多色GECIを自在に組み合わせて、単一細胞レベルで光のみで神経活動を計測中に操作するといったall-opticalイメージングが可能となる15)。特に、XCaMPセンサーの高速性を利用して、特定の頻度の神経発火時のみに、活動を操作するといったクローズドループall-opticalイメージングが可能となり、脳内における発火パターンの情報表現の因果律にまで迫ることが可能となることを大いに期待している。
本稿で紹介した研究成果は東京大学大学院医学系研究科神経生化学教室およびスタンフォード大学医学部バイオエンジニアリング専攻で得られました。多大なるご指導を賜りました尾藤晴彦教授、Karl Deisseroth教授に厚く御礼を申し上げます。特に、本プロジェクトは、一貫して、狩野方伸研究室(東大)の竹内敦也博士ならびに喜多村和郞研究室(山梨大)の真仁田聡助教との文字通り不眠不休のコラボを続けた成果であり、このような協力体制を築いて下さった両研究室に心から感謝申し上げます。また、研究遂行にあたり多大なるご助力を賜りました中井淳一研究室(埼玉大、現東北大)、根本知己研究室(北大)、横山弘之研究室(東北大)、崎村健司研究室(新潟大)、今村健志研究室(愛媛大)の皆様に深く感謝申し上げます。最後になりますが、本稿を執筆する機会を与えてくださいました日本神経化学会奨励賞選考委員の先生方、関係者の先生方に厚く御礼申し上げます。
1) Tsien RY, Pozzan T, Rink TJ. Calcium homeostasis in intact lymphocytes: Cytoplasmic free calcium monitored with a intracellularly trapped fluorescent indicator. J Cell Biol, 94(2), 325–334 (1982).
2) Miyawaki A, Llopis J, Heim R, McCaffery JM, Adams JA, Ikura M, Tsien RY. Fluorescent indicators for Ca2+ based on green fluorescent proteins and calmodulin. Nature, 388(6645), 882–887 (1997).
3) Nakai J, Ohkura M, Imoto K. A high signal-to-noise Ca(2+) probe composed of a single green fluorescent protein. Nat Biotechnol, 19(2), 137–141 (2001).
4) Tian L, Hires SA, Mao T, Huber D, Chiappe ME, Chalasani SH, Petreanu L, Akerboom J, McKinney SA, Schreiter ER, Bargmann CI, Jayaraman V, Svoboda K, Looger LL. Imaging neural activity in worms, flies and mice with improved GCaMP calcium indicators. Nat Methods, 6(12), 875–881 (2009).
5) Shindo A, Hara Y, Yamamoto TS, Ohkura M, Nakai J, Ueno N. Tissue-tissue interaction-triggered calcium elevation is required for cell polarization during Xenopus gastrulation. PLoS One, 5(2), e8897 (2010).
6) Akerboom J, Chen TW, Wardill TJ, Tian L, Marvin JS, Mutlu S, Calderón NC, Esposti F, Borghuis BG, Sun XR, Gordus A, Orger MB, Portugues R, Engert F, Macklin JJ, Filosa A, Aggarwal A, Kerr RA, Takagi R, Kracun S, Shigetomi E, Khakh BS, Baier H, Lagnado L, Wang SS, Bargmann CI, Kimmel BE, Jayaraman V, Svoboda K, Kim DS, Schreiter ER, Looger LL. Optimization of a GCaMP calcium indicator for neural activity imaging. J Neurosci, 32(40), 13819–13840 (2012).
7) Chen TW, Wardill TJ, Sun Y, Pulver SR, Renninger SL, Baohan A, Schreiter ER, Kerr RA, Orger MB, Jayaraman V, Looger LL, Svoboda K, Kim DS. Ultrasensitive fluorescent proteins for imaging neuronal activity. Nature, 499(7458), 295–300 (2013).
8) Ohkura M, Sasaki T, Sadakari J, Gengyo-Ando K, Kagawa-Nagamura Y, Kobayashi C, Ikegaya Y, Nakai J. Genetically encoded green fluorescent Ca2+ indicators with improved detectability for neuronal Ca2+ signals. PLoS One, 7(12), e51286 (2012).
9) Dana H, Sun Y, Mohar B, Hulse BK, Kerlin AM, Hasseman JP, Tsegaye G, Tsang A, Wong A, Patel R, Macklin JJ, Chen Y, Konnerth A, Jayaraman V, Looger LL, Schreiter ER, Svoboda K, Kim DS. High-performance calcium sensors for imaging activity in neuronal populations and microcompartments. Nat Methods, 16(7), 649–657 (2019).
10) Zhao Y, Araki S, Wu J, Teramoto T, Chang YF, Nakano M, Abdelfattah AS, Fujiwara M, Ishihara T, Nagai T, Campbell RE. An expanded palette of genetically encoded Ca2+ indicators. Science, 333(6051), 1888–1891 (2011).
11) Ohkura M, Sasaki T, Kobayashi C, Ikegaya Y, Nakai J. An improved genetically encoded red fluorescent Ca2+ indicator for detecting optically evoked action potentials. PLoS One, 7(7), e39933 (2012).
12) Akerboom J, Carreras Calderón N, Tian L, Wabnig S, Prigge M, Tolö J, Gordus A, Orger MB, Severi KE, Macklin JJ, Patel R, Pulver SR, Wardill TJ, Fischer E, Schüler C, Chen TW, Sarkisyan KS, Marvin JS, Bargmann CI, Kim DS, Kügler S, Lagnado L, Hegemann P, Gottschalk A, Schreiter ER, Looger LL. Genetically encoded calcium indicators for multi-color neural activity imaging and combination with optogenetics. Front Mol Neurosci, 6, 2 (2013).
13) Inoue M, Takeuchi A, Horigane S, Ohkura M, Gengyo-Ando K, Fujii H, Kamijo S, Takemoto-Kimura S, Kano M, Nakai J, Kitamura K, Bito H. Rational design of a high-affinity, fast, red calcium indicator R-CaMP2. Nat Methods, 12(1), 64–70 (2014).
14) Inoue M, Takeuchi A, Manita S, Horigane SI, Sakamoto M, Kawakami R, Yamaguchi K, Otomo K, Yokoyama H, Kim R, Yokoyama T, Takemoto-Kimura S, Abe M, Okamura M, Kondo Y, Quirin S, Ramakrishnan C, Imamura T, Sakimura K, Nemoto T, Kano M, Fujii H, Deisseroth K, Kitamura K, Bito H. Rational engineering of XCaMPs, a multicolor GECI suite for in vivo imaging of complex brain circuit dynamics. Cell, 177(5), 1346–1360.e24 (2019).
15) Marshel JH, Kim YS, Machado TA, Quirin S, Benson B, Kadmon J, Raja C, Chibukhchyan A, Ramakrishnan C, Inoue M, Shane JC, McKnight DJ, Yoshizawa S, Kato HE, Ganguli S, Deisseroth K. Cortical layer-specific critical dynamics triggering perception. Science, 365(6453), eaaw5202 (2019).

This page was created on 2019-11-13T17:47:25.463+09:00
This page was last modified on 2019-12-19T15:45:39.000+09:00
このサイトは(株)国際文献社によって運用されています。