意欲行動における腹側海馬の機能解明
慶應義塾大学医学部精神・神経科学教室
発行日:2019年12月30日Published: December 30, 2019
© 2019 日本神経化学会© 2019 The Japanese Society for Neurochemistry
意欲は、目標を達成するための行動を下支えする活力である。私達はよりよい生活を営もうと目標をもち、その目標達成に向けて行動する。この目標達成に向けた行動が意欲行動である。例えば、健康維持のために運動する、試験に合格するために勉強する、といった具合に私たちの日常生活でよくみられる行動の一つである。また、うつ病をはじめとする様々な精神疾患で意欲の障害が生じ、日常生活や社会生活に困難をきたす。ゆえに、意欲行動の神経メカニズムを知ることは、意欲行動に異常をいたす精神疾患の治療につながることが期待されるため、神経科学・精神医学の両分野を跨いだ重要なテーマとなっている。
意欲行動の背景には、目標を設定してはじめの一歩を踏み出すこと「行動の開始」と、目標達成までねばり強く行動を続けること「行動の継続」の2つがあると考えられる(図1)。特に目標が高いほど、ねばり強く行動を継続する必要がある。これまでに、はじめの一歩を踏み出す、行動を開始する脳内メカニズムについては、運動制御や報酬を計算する大脳基底核(腹側線条体やドパミン神経)が重要であることが明らかにされてきた1–3)。しかし、行動の継続について取り扱った研究がなく、その脳内メカニズムは全く分かっていなかった。本稿では、意欲行動を継続させる脳内メカニズムについて、著者らの研究成果を紹介する。
意欲行動が続けられなくなる原因として何が考えられるだろうか。その原因の一つに不安が考えられる。意欲的に取り組んでいるにもかかわらず、わずかでも不安が昂じると行動に集中できず、手を止めてしまうことは私たちの体験からも理解できる。腹側海馬は不安を制御する脳部位として知られている4)。興味深いことに、腹側海馬は腹側線条体へ密に投射することが、解剖学的研究から明らかにされている5)。これらは、意欲行動において腹側海馬の活動が重要な役割を果たすことを示唆する知見である。そこで著者らは、意欲行動の継続と腹側海馬の活動の関係に着目した。
著者らは、まず意欲行動の継続を定量する実験系をマウスで確立した。マウスには食事制限を課し、レバーを押せばエサがもらえることを事前に学習させる。テスト課題では、「制限時間」と「必要なレバー押しの回数」を設け、制限時間内にマウスが必要回数のレバーを押すことができればエサを獲得できる。設定した回数に至るまでマウスがレバーを押し続けることができず制限時間を迎えた場合、そのトライアルは失敗となる。この失敗確率により意欲行動の継続を評価した(図2)。5回のレバー押しでエサがもらえる課題での失敗確率は5%、10回のレバー押しでは27%、20回のレバー押しでは50%と、課題の難易度が増すと失敗確率が増加した。
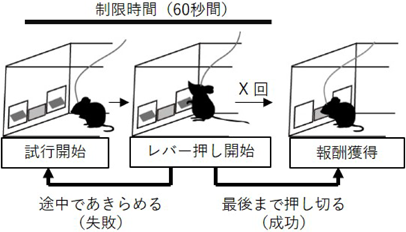
60秒間の制限時間内に実験者が設定した回数のレバーをマウスが押すことができればエサ報酬を獲得できる。失敗確率によって意欲行動の継続を評価できる課題。
腹側海馬の活動計測には、ファイバーフォトメトリー法による集団カルシウム計測系を用いた6)。海馬CA1領域特異的にカルシウム蛍光プローブを発現する遺伝子改変マウスを用い、このマウスの腹側海馬に光ファイバーを留置する(図3A)。光ファイバーを通じてプローブを励起し、神経活動によって変化する蛍光を同じ光ファイバーで回収することで、腹側海馬CA1の活動を計測することができる。この活動観察技術を用いて、レバー押し課題中に腹側海馬CA1神経細胞の活動計測を行った。その結果、試行開始から徐々に腹側海馬神経活動が下がり始め、レバー押し開始(1回目のレバー押し)からレバーを押し終わるまで腹側海馬の活動抑制が持続した(図3B)。一方、途中でレバーを押さなくなった失敗トライアルでは、腹側海馬の活動抑制が解除され、ベースラインに戻ることが分かった(図3C)。
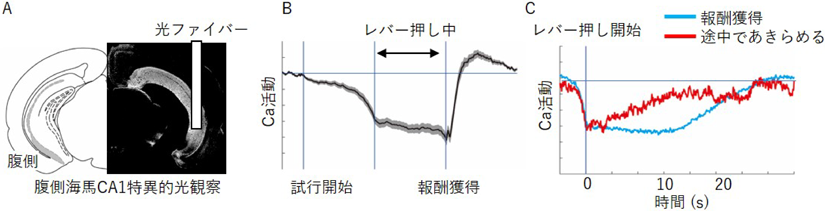
A)腹側海馬CA1へ光ファイバー刺入図。海馬CA1にカルシウム蛍光プローブが発現する遺伝子改変マウス。B)5回レバー押し課題における腹側海馬の活動。横線は活動のベースラインを示す。レバー押し行動を継続する間、腹側海馬の活動抑制が観察された。C)20回レバー押し課題における腹側海馬の活動。横線は活動のベースラインを示す。レバー押しを完遂し目標を達成できた試行では、レバー押し行動の継続中、腹側海馬の活動が抑制された(青色)。途中であきらめた試行では、腹側海馬の活動がベースラインに戻った(赤)。
意欲行動中の腹側海馬の活動観察により、意欲行動を継続している間は腹側海馬の活動が低下することが示された。では、意欲行動の継続中に腹側海馬の活動が低下することにどんな役割があるだろうか。そこで著者らは、光操作技術(オプトジェネティクス)を用いて、意欲行動の継続と腹側海馬活動の因果関係を調べた。
まず、課題中に腹側海馬を光活性化することで、意欲行動の継続を妨害するか調べた。光活性化の実験では、海馬神経細胞特異的にChR2(光活性化分子)を発現させた遺伝子改変マウスを用いた。このマウスの腹側海馬の両側に光ファイバーを留置し、レバー押し行動中に腹側海馬を光活性化した。要求されるレバー押しの数が5回の課題において、腹側海馬神経細胞を興奮させ、活動抑制を解除したところ、失敗確率が5%から20%へ増加した(図4A)。一方、課題中の行動開始時、報酬獲得後(レバー押し以外のタイミング)の光活性化では意欲行動への影響は観察されなかった。
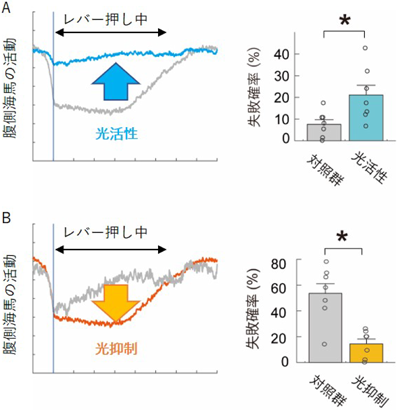
A)オプトジェネティクスで腹側海馬の活動抑制を解除すると、失敗確率が増加した(レバーを押さなくなった)。B)オプトジェネティクスで腹側海馬の活動抑制を手助けすると、失敗確率が減少した(レバーをよく押すようになった)。
次に、課題中に腹側海馬を光抑制することで、レバー押し行動を促進させるか調べた。腹側海馬の抑制には、海馬神経細胞特異的にArchT(光抑制分子)を発現させた遺伝子改変マウスを用いた。この実験では、20回のレバー押しが要求される難易度の高いオペラント課題を用いた。レバー押し行動中に腹側海馬を光抑制した結果、失敗確率が50%から17%へ減少した(図4B)。以上のことから、腹側海馬神経細胞の活動抑制が意欲行動の継続に必須であることが示唆された。
最後に著者らは、意欲行動の継続中にみられる腹側海馬神経細胞の抑制メカニズムについて探索した。正中縫線核(median raphe: MR)に局在するセロトニン神経(5-HT神経)を活性化すると、背側海馬錐体細胞の活動を抑制することが先行研究で報告されている7)。MR 5-HT神経は腹側海馬へも投射することが知られている8)。著者らはまず、MR 5-HT神経の活性化により腹側海馬の活動が低下するか検証した。この実験では、5-HT神経にChR2を発現させたマウスを用いた。MR 5-HT神経の光活性化による腹側海馬の活動応答を、電気生理学的手法を用いて計測した。照射時に、正中縫線核の背側に位置する背側縫線核(dorsal raphe: DR)を刺激しないようにするために、尾側からほぼ水平に光ファイバーを刺入し、MRをターゲットした。その結果、MR 5-HT神経の活性化により腹側海馬の神経活動が低下した。
MR 5-HT神経の活動亢進により腹側海馬の活動が低下することから、レバー押し行動中の腹側海馬の活動低下に5-HT神経活動が関与する場合、レバー押し行動中はMR 5-HT神経の活動が高いことが予想できた。そこで次に、レバー押し課題中のMR 5-HT神経の活動を計測し、意欲行動の継続中に生じる腹側海馬の活動抑制に5-HT神経が関与するかを調べた。この実験では、5-HT神経特異的にカルシウム蛍光プローブを発現する遺伝子改変マウスを用いた。その結果、マウスがレバーを押している間、MR 5-HT神経の活動亢進が観察された(図5)。
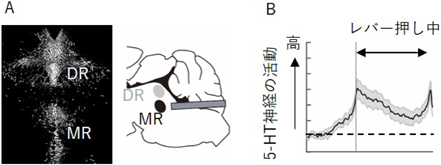
A)縫線核セロトニン神経にカルシウム蛍光プローブが発現する遺伝子改変マウス。正中縫線核(median raphe: MR)の背側に位置する背側縫線核(dorsal raphe: DR)に局在する5-HT神経からの計測を避けるために、尾側から水平に光ファイバーを刺入し、MR 5-HT神経のみをターゲットした。B)レバー押し行動中にMR 5-HT神経の活動亢進が観察された。
セロトニン受容体(Htr)には14アイソフォームが存在する9)。このうち、海馬に発現し、海馬CA1錐体細胞の活動の抑制を説明できるセロトニン受容体は、Htr1aとHtr3aの2つが考えられる10)。Htr1a(Gi共役型)は海馬CA1錐体細胞に発現する。セロトニンはGiを介してCA1錐体細胞を直接的に抑制する。一方、Htr3a(イオン透過型)は海馬GABAインターニューロンに発現する。セロトニンはHtr3a陽性インターニューロンの活動を増加させ、CA1錐体細胞を間接的に抑制する。そこで著者らは、レバー押し行動中の腹側海馬の活動抑制にどちらの候補セロトニン受容体分子が関与するか調べた。テスト課題前のマウスに対し、Htr1aもしくはHtr3a拮抗薬を腹腔内投与し、レバー押し行動を妨害するか検証した。その結果、Htr3a拮抗薬の投与によって失敗確率が増加した(図6A)。一方、Htr1a拮抗薬の投与では失敗確率への影響が見られなかった。最後に著者らは、腹側海馬に発現するHtr3aを介した抑制経路が重要であるか確認した。マウスの両側腹側海馬に薬液注入用カニューラを留置し、Htr3a拮抗薬を局所投与した結果、レバー押し課題の失敗確率が増加した。この結果から、海馬に発現するHtr3aが意欲行動の継続に関わることがわかった(図6B)。
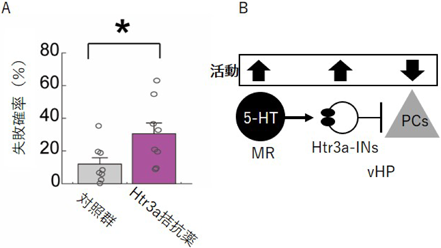
A)腹側海馬にHtr3a拮抗薬を投与すると、失敗確率が増加した。B)正中縫線核5-HT神経が活性化し、海馬で放出されるセロトニンが海馬に存在するHtr3a陽性インターニューロン(Htr3a-INs)を介して海馬神経細胞(PCs)抑制を抑制することが、意欲行動の継続に必要。
本研究により、意欲行動の継続中に腹側海馬の活動が抑制されること(I)、この活動抑制は意欲行動の継続に必要であること(II)、意欲行動の継続中に正中縫線核5-HT神経が活性化すること(III)、腹側海馬に存在するHtr3aの遮断により、意欲行動の継続を妨げること(IV)を発見した。以上の結果から、Htr3a陽性インターニューロンを介した5-HTによる腹側海馬活動の間接的な抑制が、意欲行動の継続に必要であることを明らかとした11)。
意欲の低下は、うつ病をはじめとする様々な精神・神経疾患で高頻度にみられる治療困難な症状である。意欲の低下は治療やリハビリの妨げになり、患者や支援者に悪影響をもたらす。しかし、意欲低下の神経基盤はよくわかっておらず、未だに有効な治療法は存在しません。例えば、うつ病の治療に認知行動療法があるが、これには定期的に病院に通い続ける(意欲行動の継続)が求められる。本研究により、意欲行動の継続にはセロトニンによる腹側海馬の活動抑制が必要であることを明らかにした。今後の課題として、うつ病モデルとして前臨床研究で使われている病態モデル動物を用いて、病態時の意欲低下と脳内メカニズムの関係性を調べることで、意欲的に行動が続けられなくて認知行動療法を受けることができないケースにどのような介入ができるか、という意欲低下治療の開発に貢献する研究を考えている。
本稿でご紹介させて頂いた研究遂行にあたり、多大なるご指導を受け賜りました慶應義塾大学医学部、精神・神経科学教室の三村將教授、田中謙二准教授、並びに研究室の皆様により心より感謝申し上げます。最後に、本稿執筆の機会を与えて下さいました日本神経化学会理事長の小泉修一先生、優秀賞・奨励賞選考委員会の先生方、並びに関係者の先生方に深く感謝申し上げます。
1) Phillips PEM, Stuber GD, Heien MLAV, Wightman RM, Carelli RM. Subsecond dopamine release promotes cocaine seeking. Nature, 422(6932), 614–618 (2003).
2) Cui G, Jun SB, Jin X, Pham MD, Vogel SS, Lovinger DM, Costa RM. Concurrent activation of striatal direct and indirect pathways during action initiation. Nature, 494(7436), 238–242 (2013).
3) Tsutsui-Kimura I, Natsubori A, Mori M, Kobayashi K, Drew MR, de Kerchove d’Exaerde A, Mimura M, Tanaka KF. Distinct roles of ventromedial versus ventrolateral striatal medium spiny neurons in reward-oriented behavior. Curr Biol, 27(19), 3042–3048.e4 (2017).
4) Jimenez JC, Su K, Goldberg AR, Luna VM, Biane JS, Ordek G, Zhou P, Ong SK, Wright MA, Zweifel L, Paninski L, Hen R, Kheirbek MA. Anxiety cells in a hippocampal–hypothalamic circuit. Neuron, 97(3), 670–683.e6 (2018).
5) Britt JP, Benaliouad F, McDevitt RA, Stuber GD, Wise RA, Bonci A. Synaptic and behavioral profile of multiple glutamatergic inputs to the nucleus accumbens. Neuron, 76(4), 790–803 (2012).
6) Natsubori A, Tsutsui-Kimura I, Nishida H, Bouchekioua Y, Sekiya H, Uchigashima M, Watanabe M, de Kerchove d’Exaerde A, Mimura M, Takata N, Tanaka KF. Ventrolateral striatal medium spiny neurons positively regulate food-incentive, goal-directed behavior independently of D1 and D2 selectivity. J Neurosci, 37(10), 2723–2733 (2017).
7) Varga V, Losonczy A, Zemelman BV, Borhegyi Z, Nyiri G, Domonkos A, Hangya B, Holderith N, Magee JC, Freund TF. Fast synaptic subcortical control of hippocampal circuits. Science, 326(5951), 449–453 (2009).
8) Vertes RP, Fortin WJ, Crane AM. Projections of the median raphe nucleus in the rat. J Comp Neurol, 407(4), 555–582 (1999).
9) Hannon J, Hoyer D. Molecular biology of 5-HT receptors. Behav Brain Res, 195(1), 198–213 (2008).
10) Tanaka KF, Samuels BA, Hen R. Serotonin receptor expression along the dorsal–ventral axis of mouse hippocampus. Philos Trans R Soc Lond B Biol Sci, 367(1601), 2395–2401 (2012).
11) Yoshida K, Drew MR, Mimura M, Tanaka KF. Serotonin-mediated inhibition of ventral hippocampus is required for sustained goal-directed behavior. Nat Neurosci, 22(5), 770–777 (2019).

This page was created on 2019-11-13T18:12:35.386+09:00
This page was last modified on 2019-12-16T11:56:30.000+09:00
このサイトは(株)国際文献社によって運用されています。