大脳皮質発生におけるリーリンシグナルによるニューロン移動制御
慶應義塾大学医学部解剖学教室
発行日:2020年6月30日Published: June 30, 2020
© 2020 日本神経化学会© 2020 The Japanese Society for Neurochemistry
哺乳類の大脳皮質は整然とした6層構造を持ち、高次機能の基盤となっている。この構造は発生期の高度に制御されたニューロン移動によって形成される。大脳皮質のニューロンの約70%を占める興奮性ニューロンは、胎生期の外套の脳室に面した脳室帯、あるいはそのすぐ外側にある脳室下帯で産生される。その後、ニューロンは放射状に脳表面へ向かって移動し皮質板を形成して、最終的に機能する場所へと配置する。この移動過程において、ニューロンは複数の異なる移動様式を示し、これらの移動が障害されると、滑脳症などの神経疾患を引き起こす。ニューロン移動と層構造は多数のシグナル経路によって制御されるが、リーリンシグナルはその主要なシグナルの一つである。巨大細胞外タンパク質であるリーリンは主に大脳皮質の表層にある辺縁帯に含まれるカハールレチウス細胞から分泌され、移動中のニューロンに作用する。本稿ではリーリンシグナルによるニューロン移動の制御機構を概説するとともに、筆者らが最近報告したニューロン移動最終段階におけるリーリン受容体の機能について紹介する。
形成中の大脳皮質において、新生ニューロンは複数の移動様式を示す。最も早く生まれたニューロンは脳室帯の外側にプレプレートと呼ばれる分化したニューロンから成る層を形成する。続いて脳室帯で生じたニューロンはプレプレートに割って入り、プレプレートは表層側の辺縁帯と深層側のサブプレートに分かれる。そして続いて移動してきた新生ニューロンはサブプレートを通り抜け、辺縁帯の直下で停止することにより皮質板を形成する。皮質形成の早期では、将来深層ニューロンになる早生まれのニューロンは先導突起を辺縁帯まで伸長させたのちに、先導突起を短くすることで細胞体を最終的な配置へと移動させる(図1)。この移動様式ではニューロンは放射状グリア非依存的に移動を行い、ソーマルトランスロケーションと呼ばれる1)。皮質形成の後期になると、皮質は厚みを増しニューロンは複数の移動様式を示すようになる。まず新生ニューロンは脳室帯で針状の形態をとり、次に脳室帯から脳表側へ出て多極性細胞となる。多極性細胞は脳室帯直上に約24時間留まり、この領域は多極性細胞蓄積帯(multipolar accumulation zone; MAZ)と呼ばれる。MAZではニューロンは多極性細胞移動と呼ばれるユニークな移動様式を示す。すなわち、ニューロンはダイナミックに多数の突起を伸縮させ、細胞体は漂うように移動する2, 3)。そののち、多極性細胞は双極性へと形態を変化させ、中間帯・皮質板で放射状グリアの突起を足場として方向性を持った移動を開始する。この放射状グリア依存的な移動はロコモーションと呼ばれる1)。ロコモーションを行うニューロンは脳表層へ向かって移動し、皮質板の最表層の細胞密度の高い領域の付近へ到達する。この領域は未成熟のニューロンの層であり、primitive cortical zone(PCZ)と呼ばれる。PCZ付近に到達したニューロンは一時的に移動を停止し、移動様式をターミナルトランスロケーションへと変化させる4)。ターミナルトランスロケーションとソーマルトランスロケーションには形態的に類似点がみられる。すなわちニューロンは脳膜側に伸ばした突起を縮めながら、細胞体を放射状グリア非依存的に急速に外側に移動させ、辺縁帯直下に到達して移動を完了する1)。これらのプロセスの中で、遅生まれのニューロンは皮質板で早生まれのニューロンを通り越して皮質板最外側部まで到達することにより、誕生時期依存的な“inside-out”パターンを形成する。つまり、遅生まれのニューロンは表層へと配置し、早生まれのニューロンは深層へと配置する。この“inside-out”パターンは哺乳類に特異的にみられることから、大脳新皮質の拡大と高度に複雑化した哺乳類の脳の進化の過程に重要であると考えられる。
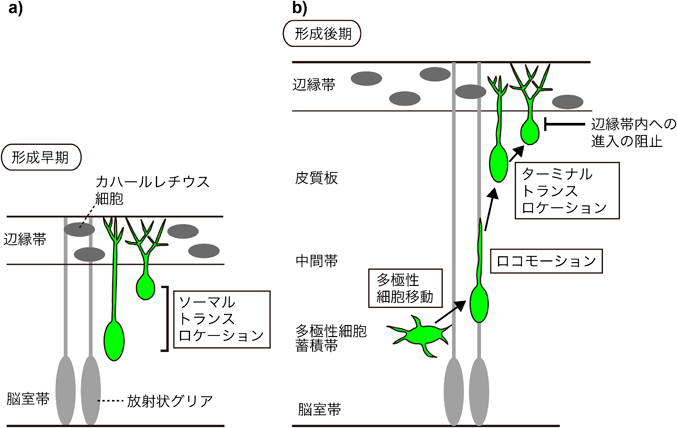
細胞外糖タンパク質であるリーリンは胎生期のニューロン移動と層形成に重要な役割を担うことが知られている。リーリンは歩行の際によろめく表現型を示す自然発症劣性変異体マウスであるリーラーの原因遺伝子として同定された5–7)。リーラーマウスの中枢神経系では大脳皮質の層構造の異常を含めて多くの異常が認められる。誕生期の細胞を標識すると、大脳皮質のニューロンの配置は概ね逆転していることからリーリンがニューロン移動と“inside-out”パターンの層形成に重要な機能を持つと考えられた。リーリンは進化的に保存された全長約3500アミノ酸残基からなる巨大な細胞外糖タンパク質であり、その主要な受容体としてリポタンパク質受容体であるapolipoprotein E receptor 2(ApoER2)とvery low density lipoprotein receptor(VLDLR)が同定された8, 9)。2種類の受容体のうち一方が損なわれたノックアウトマウスでは層構造の異常は軽度だが、ダブルノックアウトマウスではリーラーマウスと同様の層構造の逆転がみられることから、これらの受容体は重複した機能を持っていると考えられている10)。リーリンが受容体に結合すると、受容体の細胞内ドメインに結合する細胞内エフェクターDab1のリン酸化が引き起こされる。このリン酸化はSrcファミリーキナーゼであるFynとSrcによって行われる11)。Dab1のノックアウトマウスに加えて、FynとSrcのダブルノックアウトマウス、Dab1のチロシンリン酸化サイトの全てに変異を持つ変異マウスも、リーラーマウスと同様の層構造異常を示すことから、Dab1のリン酸化がリーリンシグナルに必須であると考えられている。リン酸化されたDab1はCrk/CrkL、SOCS3、Nckβ、PI3K、Lis1等の多様な分子と相互作用し、下流経路にシグナルを伝達する。
発生中の大脳皮質において、リーリンは主に辺縁帯に含まれるカハールレチウス細胞から分泌される6, 12)。さらに、少量のリーリンは中間帯の深部にも検出される13–15)。またリーリンは蛋白質分解を受けて小断片となり、皮質板の深部へと拡散することも示されている16)。リーリン受容体のうちApoER2はMAZおよび中間帯の深部に豊富に発現しており13, 14)、アルカリフォスファターゼとリーリンの受容体結合部位の融合蛋白質を用いて、リーリンが実際にこの部位で受容体に結合することも示されている14)。この知見に一致して、ApoER2ノックアウトマウスおよびリーラーマウスでは中間帯に移動ニューロンが停滞する17–19)。この機能を担う下流経路の一つとして、Nカドヘリンを介した細胞接着の制御が考えられる。ドミナントネガティブ型のVLDLRを発現させてリーリンと受容体の結合を阻害すると、Nカドヘリンを介した多極性細胞から双極性細胞への変換が損なわれることが示された20)。また、リーリンシグナルはLIMK1の活性化を介してアクチン脱重合因子cofilinの活性を上昇させる21)。リーラーマウスにおける放射状移動の異常が、野生型LIMK1あるいはcofilinのphosphomimic型変異体を発現させることによりレスキューされることから、リーリンシグナル下流でのアクチンの制御も中間帯での放射状移動に関与すると考えられている22)。
ニューロンは脳表層にある辺縁帯の下部に到達すると移動を終える。辺縁帯にはリーリンが高濃度に存在し、受容体の発現も認められることからリーリンシグナルがこのタイミングでニューロンに作用することが推定された10, 14, 19, 23)。この推定に一致して、リーリンシグナルが辺縁帯直下で、細胞接着因子であるインテグリンα5β1を活性化し、ニューロンの樹状突起を辺縁帯にある細胞外マトリックスであるフィブロネクチンに係留させることによりターミナルトランスロケーションを促進することが示された24)。また前述のリーリンシグナルによるLIMK1/cofilinを介したアクチン細胞骨格の制御も、ニューロンの先導突起を安定化することによりターミナルトランスロケーションを促進することが示された21, 25)。
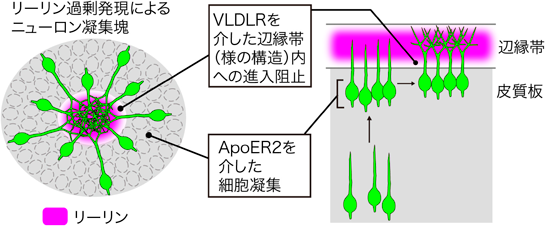
ターミナルトランスロケーション後のニューロンでは、細胞体は辺縁帯の直下に留まり、辺縁帯内部に進入することはない。VLDLRノックアウトマウスにおいて辺縁帯内部のニューロンの数の増加が示されていたが、胎仔期のニューロン移動の異常に起因するものかは不明だった18)。筆者らは、脳室帯で産生されたニューロンを子宮内電気穿孔法によってGFPで標識しその移動分布を観察する方法により、リーリン受容体のノックアウトマウスでは、胎仔期に辺縁帯内部へと少数のニューロンが進入することを明らかにした19, 26)。ApoER2ノックアウトマウスでは前述のように中間帯での放射状移動に異常があるため、その後のニューロン移動にも二次的に影響を与える可能性があるのに対して、VLDLRノックアウトマウスでは放射状移動には異常がみられないことから、辺縁帯付近でのリーリンシグナルの機能を調べるためにはVLDLRノックアウトマウスが有用であると考えられた。VLDLRノックアウトマウスでは早生まれおよび遅生まれのニューロンとも、皮質板外側部に到達したニューロンの分布がコントロールに比べて表層寄りへとシフトしていたことから、リーリンシグナルがVLDLRを介してニューロンの過剰な移動を抑制し、辺縁帯への進入を阻止していると考えられた。VLDLRノックアウトマウスで認められた辺縁帯内へのニューロンの進入は、インテグリンα5とセリンスレオニンリン酸化酵素Aktおよび恒常的活性型Rap1の導入により回復がみられた。またリーリンシグナルは大脳皮質、海馬の形成において神経突起の伸長を促進することが報告されており、VLDLRノックアウトマウスにおいても辺縁帯内部に伸長している樹状突起の低形成が認められた。これらの結果から、リーリンシグナルは移動を終了しつつある神経細胞において、樹状突起伸長および辺縁帯内部への進入の阻止を制御すると考えられる。
野生型マウス胎仔期の大脳皮質に異所的にリーリンを発現させると移動中のニューロンが細胞凝集塊を形成する23)。この際、ニューロンは細胞凝集塊中に分泌されたリーリンに沿って移動を停止し、正常な発生過程と類似した誕生時期依存的な“inside-out”パターンの配置をする。またこの細胞塊には中央部分に細胞密度が低い領域があり、その周囲にニューロンの細胞体が高密度に分布しており、生体内で観察される辺縁帯と類似しているため、リーリンは移動中のニューロンに作用して辺縁帯様の構造を作り出す能力を持つと考えられる。リーリン受容体のノックアウトマウスでリーリンを発現させると、ApoER2ノックアウトマウスでは細胞凝集塊の形成が認められず、VLDLRノックアウトマウスでは細胞凝集塊は形成されるが、野生型の細胞凝集塊の中央でみられる辺縁帯様の中空構造は観察されなかった19, 26)。この結果は、ニューロンの凝集過程にはApoER2が必要であるがVLDLRは寄与せず、細胞が凝集したあとの樹状突起と細胞体を分離させる過程にVLDLRが必要であることを示唆している。よって2種類のリーリン受容体はニューロン移動の最終過程において異なる機能を果たしていると考えられる(図2)。
これまでの筆者らの研究により、VLDLRはニューロンが移動を終え、正常な配向を形成する過程に機能すると考えられる。ではVLDLRはどのように辺縁帯内部へのニューロン進入を阻止しているのだろうか。近年、ニューロン移動後の配置と神経突起の形成の関連が複数の系で示されている。筆者らの研究ではVLDLRが樹状突起の形成に重要であることが示された。またインテグリンα5の関与が示されたことを合わせて考えると、VLDLRが関与する樹状突起形成と細胞接着を介して、樹状突起と細胞体の分離は制御されている可能性が考えられる。今後、2種類のリーリン受容体が異なる機能を発揮するメカニズムを明らかにすることで、大脳皮質層形成におけるリーリンシグナルの寄与の全貌に迫りたい。
本稿で紹介した研究は慶應義塾大学医学部解剖学教室で仲嶋一範先生のご指導のもと、教室員の皆様と共同研究者の方々のご協力を得て行ったものであり、皆様に深く感謝申し上げます。また、今回執筆の機会を与えてくださいました神経化学会出版・広報委員会、関係者の皆様に感謝致します。
1) Nadarajah B, Brunstrom JE, Grutzendler J, Wong RO, Pearlman AL. Two modes of radial migration in early development of the cerebral cortex. Nat Neurosci, 4(2), 143–150 (2001).
2) Noctor SC, Martinez-Cerdeno V, Ivic L, Kriegstein AR. Cortical neurons arise in symmetric and asymmetric division zones and migrate through specific phases. Nat Neurosci, 7(2), 136–144 (2004).
3) Tabata H, Nakajima K. Multipolar migration: The third mode of radial neuronal migration in the developing cerebral cortex. J Neurosci, 23(31), 9996–10001 (2003).
4) Sekine K, Honda T, Kawauchi T, Kubo K, Nakajima K. The outermost region of the developing cortical plate is crucial for both the switch of the radial migration mode and the dab1-dependent “inside-out” lamination in the neocortex. J Neurosci, 31(25), 9426–9439 (2011).
5) Bar I, Lambert De Rouvroit C, Royaux I, Krizman DB, Dernoncourt C, Ruelle D, Beckers MC, Goffinet AM. A YAC contig containing the reeler locus with preliminary characterization of candidate gene fragments. Genomics, 26(3), 543–549 (1995).
6) D’Arcangelo G, Miao GG, Chen SC, Soares HD, Morgan JI, Curran T. A protein related to extracellular matrix proteins deleted in the mouse mutant reeler. Nature, 374(6524), 719–723 (1995).
7) Hirotsune S, Takahara T, Sasaki N, Hirose K, Yoshiki A, Ohashi T, Kusakabe M, Murakami Y, Muramatsu M, Watanabe S, Nakao K, Katsuki M, Hayashizaki Y. The reeler gene encodes a protein with an EGF-like motif expressed by pioneer neurons. Nat Genet, 10(1), 77–83 (1995).
8) D’Arcangelo G, Homayouni R, Keshvara L, Rice DS, Sheldon M, Curran T. Reelin is a ligand for lipoprotein receptors. Neuron, 24(2), 471–479 (1999).
9) Hiesberger T, Trommsdorff M, Howell BW, Goffinet A, Mumby MC, Cooper JA, Herz J. Direct binding of Reelin to VLDL receptor and ApoE receptor 2 induces tyrosine phosphorylation of disabled-1 and modulates tau phosphorylation. Neuron, 24(2), 481–489 (1999).
10) Trommsdorff M, Gotthardt M, Hiesberger T, Shelton J, Stockinger W, Nimpf J, Hammer RE, Richardson JA, Herz J. Reeler/Disabled-like disruption of neuronal migration in knockout mice lacking the VLDL receptor and ApoE receptor 2. Cell, 97(6), 689–701 (1999).
11) Howell BW, Herrick TM, Cooper JA. Reelin-induced tyrosine [corrected] phosphorylation of disabled 1 during neuronal positioning. Genes Dev, 13(6), 643–648 (1999).
12) Ogawa M, Miyata T, Nakajima K, Yagyu K, Seike M, Ikenaka K, Yamamoto H, Mikoshiba K. The reeler gene-associated antigen on Cajal–Retzius neurons is a crucial molecule for laminar organization of cortical neurons. Neuron, 14(5), 899–912 (1995).
13) Hirota Y, Kubo K, Katayama K, Honda T, Fujino T, Yamamoto TT, Nakajima K. Reelin receptors ApoER2 and VLDLR are expressed in distinct spatiotemporal patterns in developing mouse cerebral cortex. J Comp Neurol, 523(3), 463–478 (2015).
14) Uchida T, Baba A, Perez-Martinez FJ, Hibi T, Miyata T, Luque JM, Nakajima K, Hattori M. Downregulation of functional Reelin receptors in projection neurons implies that primary Reelin action occurs at early/premigratory stages. J Neurosci, 29(34), 10653–10662 (2009).
15) Yoshida M, Assimacopoulos S, Jones KR, Grove EA. Massive loss of Cajal–Retzius cells does not disrupt neocortical layer order. Development, 133(3), 537–545 (2006).
16) Jossin Y, Gui L, Goffinet AM. Processing of Reelin by embryonic neurons is important for function in tissue but not in dissociated cultured neurons. J Neurosci, 27(16), 4243–4252 (2007).
17) Britto JM, Tait KJ, Johnston LA, Hammond VE, Kalloniatis M, Tan SS. Altered speeds and trajectories of neurons migrating in the ventricular and subventricular zones of the reeler neocortex. Cereb Cortex, 21(5), 1018–1027 (2011).
18) Hack I, Hellwig S, Junghans D, Brunne B, Bock HH, Zhao S, Frotscher M. Divergent roles of ApoER2 and Vldlr in the migration of cortical neurons. Development, 134(21), 3883–3891 (2007).
19) Hirota Y, Kubo KI, Fujino T, Yamamoto TT, Nakajima K. ApoER2 controls not only neuronal migration in the intermediate zone but also termination of migration in the developing cerebral cortex. Cereb Cortex, 28(1), 223–235 (2018).
20) Jossin Y, Cooper JA. Reelin, Rap1 and N-cadherin orient the migration of multipolar neurons in the developing neocortex. Nat Neurosci, 14(6), 697–703 (2011).
21) Chai X, Forster E, Zhao S, Bock HH, Frotscher M. Reelin stabilizes the actin cytoskeleton of neuronal processes by inducing n-cofilin phosphorylation at serine3. J Neurosci, 29(1), 288–299 (2009).
22) Chai X, Zhao S, Fan L, Zhang W, Lu X, Shao H, Wang S, Song L, Failla AV, Zobiak B, Mannherz HG, Frotscher M. Reelin and cofilin cooperate during the migration of cortical neurons: A quantitative morphological analysis. Development, 143(6), 1029–1040 (2016).
23) Kubo K, Honda T, Tomita K, Sekine K, Ishii K, Uto A, Kobayashi K, Tabata H, Nakajima K. Ectopic Reelin induces neuronal aggregation with a normal birthdate-dependent “inside-out” alignment in the developing neocortex. J Neurosci, 30(33), 10953–10966 (2010).
24) Sekine K, Kawauchi T, Kubo K, Honda T, Herz J, Hattori M, Kinashi T, Nakajima K. Reelin controls neuronal positioning by promoting cell-matrix adhesion via inside-out activation of integrin alpha5beta1. Neuron, 76(2), 353–369 (2012).
25) Franco SJ, Martinez-Garay I, Gil-Sanz C, Harkins-Perry SR, Muller U. Reelin regulates cadherin function via Dab1/Rap1 to control neuronal migration and lamination in the neocortex. Neuron, 69(3), 482–497 (2011).
26) Hirota Y, Nakajima K. VLDLR is not essential for Reelin-induced neuronal aggregation but suppresses neuronal invasion into the marginal zone. Development, 147(12), dev189936 (2020).

This page was created on 2020-05-26T09:09:00.119+09:00
This page was last modified on 2020-08-24T10:54:50.000+09:00
このサイトは(株)国際文献社によって運用されています。