「脳の窓」脳室周囲器官の動的な血管構築から読み解く脳と全身の情報交換機構
奈良県立医科大学・医学部・解剖学第二講座
発行日:2020年12月30日Published: December 30, 2020
脳室周囲器官は血液脳関門を持たない脳部位の総称である。血液脳関門を持たないことで、血中の分子を感知し、脳で産生したホルモンを血中へ分泌するため、「脳の窓」と呼ばれる。しかし、血液脳関門を持たないがゆえに、神経細胞が血中の有害な分子にさらされる可能性もある。著者らは①脳室周囲器官の血管はどのようなメカニズムで血液脳関門を持たないのか。②血液脳関門を持たない脳室周囲器官の神経組織はどのように保護されているのか。③脳室周囲器官の一つ脳弓下器官は血液中の細菌菌体成分リポ多糖(Lipopolysaccharides: LPS)をどのように感知しているのか。という3つの問いに取り組み、①血管のリモデリングが起きることで血管透過性・可塑性を高く保っている。②血管が動的である代わりにアストロサイトによるグリア瘢痕様構造と、二重の血管基底膜からなる血管周囲腔が血中物質の透過を制限している。③血管周囲腔に存在する単球-マクロファージ系細胞が末梢由来LPSを感知して炎症性サイトカインインターロイキン-1β(interleukin-1β: IL-1β)を産生するが、エンドトキシン耐性マウスではそのIL-1β産生が減衰する。ということを明らかにした(図1)。

© 2020 日本神経化学会© 2020 The Japanese Society for Neurochemistry
血中に投与した水銀が神経組織に透過せずに血管内にとどまることを、1695年発行の著書に記したRidley博士が、血液脳関門の発見者であるとされている1)。その後Ehrlich博士とGoldmann博士が生体染色色素であるトリパンブルーを動物に末梢から投与すると、動物の内臓は青く染まるが脳や脊髄はほとんど染色されないことを報告し、この実験から血液脳関門の存在が知られるようになった2–4)。その後血液脳関門は、血漿成分内の神経毒性物質、血球、病原体の自由な侵入を防ぐことにより神経組織内環境を保護することが分かった。血液脳関門の破綻は、血液由来物質の神経組織への透過と神経損傷をもたらす。
脳の中にも、例外的に末梢から投与したトリパンブルーに染色される、つまり血液脳関門を持たない脳部位が複数存在し、それらを総称して脳室周囲器官という。脳室周囲器官の血管は、血液脳関門を持たないことで血液循環を介した脳と全身の間の情報交換を担うため、「脳の窓」と呼ばれる5)。これは、日本人になじみ深い言葉で言えば、鎖国を敷いた江戸時代の「出島」のようなものである。脳室周囲器官は体液調節・発熱・食欲調節・睡眠・悪心嘔吐・神経内分泌といった生命維持に必要な脳機能に広く関与することがこれまでに示されている。ではどのようなメカニズムで脳室周囲器官は血液脳関門を持たないのか。また血液脳関門を持たないで脳室周囲器官の神経組織はどのように保護され、かつ物質の往来を可能にしているのであろうか。本稿では、これらの問いに取り組んできた著者らのこれまでの研究成果を中心に概説する。
脳室周囲器官には、終板器官、脳弓下器官、正中隆起、下垂体後葉、最後野、松果体が含まれ、いずれも第3脳室または第4脳室の周囲に存在する。終板器官は血漿Na+イオンと浸透圧を感知することにより、体液の恒常性に重要な役割を果たす。脳弓下器官は血漿及び脳脊髄液中のNa+レベル及びアンジオテンシンIIの増加に応答することが示されている。最後野は催吐剤に刺激される延髄嘔吐中枢の一部であり、心血管及び呼吸の調節にも関与している。これら三つの脳室周囲器官は、血中物質を感知する役割を持つので感知系脳室周囲器官と呼ばれ、各種血中物質の受容体が存在し脳室周囲器官の中でも神経細胞の細胞体が豊富に存在する部位である6, 7)。神経軸索終末からオキシトシンとアルギニン-バゾプレッシンが血中に分泌される下垂体後葉、下垂体前葉ホルモン放出ホルモンが分泌される正中隆起、メラトニンが分泌される松果体を合わせて分泌系脳室周囲器官と呼ぶ。しかし、正中隆起は近傍の視床下部弓状核へレプチンやインスリンをはじめとした代謝シグナルを伝える、つまり感知機能の一部も担っているため、一概に感知系・分泌系と二分できるわけではない。脳脊髄液を産生する脈絡叢は、有窓性の毛細血管を持つため脳室周囲器官に含まれることもあるが、神経組織を持たない。交連下器官も脳室周囲器官に含む研究者もいるが、有窓性の毛細血管は持たない。また脳室周囲器官は感染性病原体が脳に侵入する入り口であるという報告もあり、プリオン病、アフリカ・トリパノソーマ症、多発性硬化症などへの関与が示唆されている8, 9)。
著者らは、脳室周囲器官研究を始める前に細胞外マトリックスの構成成分であるコンドロイチン硫酸プロテオグリカンの発現分布を調べていて、下垂体後葉の血管にその発現を認めた10)。研究を進めるうちに、NG2プロテオグリカンの発現が下垂体後葉を含む脳室周囲器官のペリサイトで共通して高いことがわかった。NG2プロテオグリカンは、未熟または活性化したペリサイトに高い発現を示し、一般的に成熟脳血管では発現が低い。つまり、脳室周囲器官では成体においても常に血管新生が起きていて、それが血液脳関門の形成を妨げているのではないかと考え、この仮説の検証を行った。脳室周囲器官では、NG2プロテオグリカンだけでなく血管新生を制御する血管内皮成長因子(vascular endothelial growth factor-A: VEGF-A)とその受容体である血管内皮成長因子受容体2(vascular endothelial growth factor receptor 2: VEGFR2)をはじめとして、低酸素誘導因子(hypoxia Inducible Factor-α: HIF1-α)、血小板由来成長因子(platelet-derived growth factor-B: PDGF-B)、PDGF-Bの受容体である血小板由来成長因子受容体β(platelet derived growth factor receptorβ: PDGFRβ)といった血管新生に関与する因子の発現が高く、血管内皮細胞のfilopodiaや増殖が認められた。血管内皮細胞の細胞死も起きていたことから、常時血管のリモデリングがなされていることが示唆される。VEGF-A受容体阻害剤や細胞増殖阻害剤(1-β-D-arabinofuranosyl cytosine: AraC)の投与により内皮細胞の増殖、微小血管の面積や直径の減少、血管透過性低下が認められたことから、脳室周囲器官では血管新生により血管透過性が高く保たれていると考えられる11–14)(図2)。また、マウスに水の代わりに塩水を飲ませると、NG2やPDGFRβの発現が増加し血管透過性が上昇した13, 15)。一方LPS投与により血管透過性と内皮細胞増殖が低下した16)。絶食でVEGF-A依存的に正中隆起の血管透過性が上がることも報告された17)。以上の結果から、脳室周囲器官では血管が動的で環境変化に応じて血管透過性を変えていることがわかった。
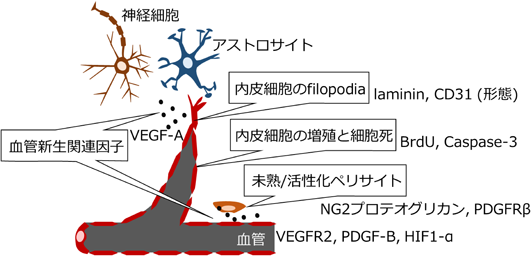
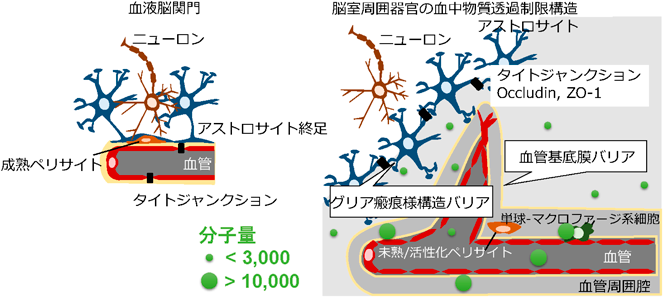
成体脳では血液脳関門を形成することで血中の有害な物質から神経細胞を保護しているが、血液脳関門を持たない脳室周囲器官の神経細胞はどのように保護されているのか?一般的な血液脳関門では内皮細胞間にタイトジャンクションを形成し、成熟したペリサイトが内皮細胞を被覆する。これをアストロサイトの終足が取り囲み、血管内皮細胞由来とアストロサイト由来の二重の血管基底膜が密着して血管内皮細胞を取り巻いている。一方、脳室周囲器官の血管は内皮細胞間のタイトジャンクションを欠いている。前項でペリサイトは未熟、または活性化していることを記した。アストロサイトは血管を取り囲むようには存在せず、二重の血管基底膜の間には大きな血管周囲腔が認められる。このように全く異なる血管構築を成すことから、脳室周囲器官には血液脳関門に代わるバリアがあるのではないかと考えた。血液脳関門の代わりのバリアを探索するに当たり、様々な分子量のトレーサーを末梢から投与して脳室周囲器官における分布を調べた。
まず分子量3,000以下の比較的小さい分子の血管透過性を評価した。フルオレセインやエバンスブルーは低分子量分子で、広く血管透過性評価に用いられているが、多くの蛍光トレーサーは血管内皮細胞に取り込まれるので神経組織に漏出しているかどうか免疫組織化学で内皮細胞を可視化して確かめる必要がある。ところが、灌流固定などのサンプル作製の過程で拡散してしまうという欠点がある。著者らはフルオレセインイソチオシアネート(fluorescein isothiocyanate: FITC)を使用して低分子量物質の血管透過性を調べるための新しい手法を開発した18)。FITCは分子量390の低分子量分子で、細胞成分の第一級アミン基に共有結合するので、免疫組織化学と組み合わせて神経組織への漏出を可視化できる。FITCの血管透過性は神経細胞の細胞体が豊富な終板器官や脳弓下器官、最後野と比較して、神経細胞の細胞体が少ないまたは存在しない正中隆起や下垂体後葉で高いことが分かった19)。また、リジン固定可能で免疫染色に耐えるデキストラン3,000も脳室周囲器官で高い透過性を示した20, 21)。さらに、脳室周囲器官のアストロサイトに着目するとグリア瘢痕様の密度の高い複雑な構造を成し、通常血液脳関門に存在するタイトジャンクションタンパク(occludin, ZO-1)を発現していた。低分子量トレーサーの神経組織への漏出は、このグリア瘢痕様構造と血管の間にのみ認められた。デキストラン10,000(分子量10,000)、デキストラン70,000(分子量70,000)、西洋わさびペルオキシダーゼ(分子量40,000)といった大きな分子は脳室周囲器官の血管周囲に限局する22)。脳室周囲器官には厚い二重の基底膜からなる血管周囲腔が存在するが、大きい分子(分子量10,000以上)は終板器官、脳弓下器官、最後野、正中隆起において血管周囲腔に集積した20, 23)。さらに、血管周囲に集積したトレーサーの血管周囲腔に存在する単球-マクロファージ系細胞への取り込みが認められた25)。
つまり、脳室周囲器官では血管透過性が高い代わりに、グリア瘢痕様構造や二重の基底膜からなる血管周囲腔により物質透過を制限し、神経細胞を保護していることが示唆された20)(図3)。正中隆起の血管構築と透過性については総説にまとめたのでご覧いただければ幸甚である27)。
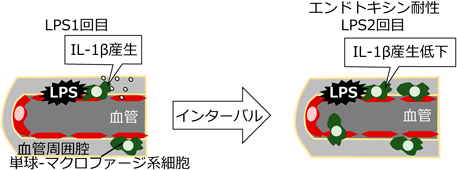
さらに、脳室周囲器官の実際の血中物質の感知機構に迫った。脳室周囲器官の一つ脳弓下器官は、細菌の菌体成分であるLPSを感知し、いち早く炎症性サイトカインIL-1βを産生する。しかし、前項の結果から、みかけの分子量が300,000以上にもなるミセルを形成するLPSは、脳弓下器官の血管周囲に留まり、脳で炎症性サイトカインを産生するミクログリアには直接アクセスできないと考えられた。LPSを投与したマウスでは発熱や自発活動量・摂食量・飲水量の低下といった行動変化が認められるが、LPS再投与時にこれらの反応が減衰するエンドトキシン耐性とよばれる現象が知られる。これはエンドトキシンの過剰反応から宿主を保護する重要なシステムであるが、そのメカニズムは不明であった。そこで著者らは、脳弓下器官においてLPSの感知機構とIL-1βの産生細胞について調べた25)。末梢から投与したLPSは血液を介して脳弓下器官に到達したが、血管周囲腔に限局した。脳で炎症性サイトカインを産生するのは一般的にはミクログリアだが、LPSは直接神経組織のミクログリアにアクセスできないと考えられる。LPSの投与後脳弓下器官でIL-1βを産生したのは血管周囲腔のIba1陽性細胞であり、この細胞が末梢由来の単球-マクロファージ系細胞であることを骨髄移植実験により明らかにした。クロドロン酸を用いて単球-マクロファージ系細胞を除去すると、LPS投与後の脳弓下器官におけるIL-1β発現が減衰した。LPSを前投与したエンドトキシン耐性マウスでは、2回目に投与したLPSも脳弓下器官の血管周囲腔に到達しているにも関わらず血管周囲の単球-マクロファージ系細胞によるIL-1β発現が著しく減衰した。これらの結果は脳弓下器官では血管周囲腔の単球-マクロファージ系細胞が血中LPSに反応してIL-1βを産生する細胞であり、単球-マクロファージ系細胞の血中LPSに対する反応性が低下することでエンドトキシン耐性が生じることを示唆する(図4)。
一般的な成体の脳血管では、損傷や低酸素脳症等の病的な状態を除いて血管新生が起きないとされてきた。本研究は①正常成体脳の中でも脳室周囲器官では定常的に血管リモデリングが起きていることを初めて報告したものであり、血管新生が脳室周囲器官の血管構造と透過性の維持に必要であることを示した。②グリア瘢痕様構造と血管周囲腔による物質透過制限は動的血管の存在による神経組織の脆弱性を補うと考えられた。③血管周囲には末梢血液由来の単球-マクロファージ系細胞が存在し、末梢血液由来LPSに応答してIL-1βを産生した。単球-マクロファージ系細胞がLPS感知を担い、神経組織の神経細胞やミクログリアがLPSを直接受容するわけではないと考えられる。加えて単球-マクロファージ系細胞の応答性が下がることがマウスの個体レベルでのエンドトキシン耐性の原因である可能性を示した。
著者は研究を通じて、人々の生活に身近でつらい症状とされる悪心嘔吐・不眠・食欲不振・過食/拒食・やる気が出ない等の緩和に貢献したい。多くの人がこれらの症状を経験しているにもかかわらず機序に不明な点が多い。機序解明の手段として、血液を介した脳と全身の情報交換機構を明らかにすることを研究のターゲットとし、新たな知見が少しでも役に立てば嬉しい。

この度は名誉ある日本神経化学会奨励賞を賜り、優秀賞・奨励賞選考委員会の先生方、学会関係者の先生方に厚く御礼申し上げます。また、本稿執筆の機会を与えてくださいましたことに感謝申し上げます。思い返せば、研究室に出入りしはじめて間もない2009年、初めて連れて行っていただいた学会が日本神経化学会でした。まだ右も左もわかりませんでしたが、活発に議論されている先生方を拝見し、いつか私もあの場に立ちたいと思いました。2015年には初めて口演を行い、たくさんのご質問、激励のお言葉とアドバイスを下さり、未熟なものを育てようとしてくださる学会のスタンスを知りました。この受賞は現時点では身に余るものですが、背伸びを許してくださる学会の先生方の胸を借り、今後の成長の糧にしたいです。
本稿でご紹介させていただいた研究遂行にあたり、多大なるご指導を賜りました京都工芸繊維大学の宮田清司教授、奈良県立医科大学医学部の和中明生教授、並びに研究室の皆様に心より感謝申し上げます。骨髄移植実験を教えてくださった杏林大学の石井さなえ先生にも感謝申し上げます。
1) Ridley H. The anatomy of the brain. London: Sam Smith and Benjamin Walford, Printers to the Royal Society (1695).
2) Mott FW. The Late Professor Edwin Goldmann’s investigations on the central nervous system by vital staining. BMJ, 2(2753), 871–873 (1913).
3) Ehrlich P. Sauerstoff-Bedürfniss des Organismus Eine farbenanalytische Studie. Berlin (1885).
4) Goldmann EE. Die äussere und Sekretion des gesunden und kranken Organismus im Lichte der “vitalen Färbung”. Beitr Klin Chirurz, 64, 192–265 (1909).
5) Gross PM, Weindl A, Knigge KM. Peering through the windows of the brain. J Cereb Blood Flow Metab, 7(6), 663–672 (1987).
6) Nakano Y, Furube E, Morita S, Wanaka A, Nakashima T, Miyata S. Astrocytic TLR4 expression and LPS-induced nuclear translocation of STAT3 in the sensory circumventricular organs of adult mouse brain. J Neuroimmunol, 278, 144–158 (2015).
7) Mannari T, Morita S, Furube E, Tominaga M, Miyata S. Astrocytic TRPV1 ion channels detect blood-borne signals in the sensory circumventricular organs of adult mouse brains. Glia, 61(6), 957–971 (2013).
8) Siso S, Gonzalez L, Jeffrey M. Neuroinvasion in prion diseases: The roles of ascending neural infection and blood dissemination. Interdiscip Perspect Infect Dis, 2010, 747892 (2010).
9) Siso S, Jeffrey M, Gonzalez L. Sensory circumventricular organs in health and disease. Acta Neuropathol, 120(6), 689–705 (2010).
10) Morita S, Oohira A, Miyata S. Activity-dependent remodeling of chondroitin sulfate proteoglycans extracellular matrix in the hypothalamo-neurohypophysial system. Neuroscience, 166(4), 1068–1082 (2010).
11) Morita S, Furube E, Mannari T, Okuda H, Tatsumi K, Wanaka A, Miyata S. Vascular endothelial growth factor-dependent angiogenesis and dynamic vascular plasticity in the sensory circumventricular organs of adult mouse brain. Cell Tissue Res, 359(3), 865–884 (2015).
12) Furube E, Mannari T, Morita S, Nishikawa K, Yoshida A, Itoh M, Miyata S. VEGF-dependent and PDGF-dependent dynamic neurovascular reconstruction in the neurohypophysis of adult mice. J Endocrinol, 222(1), 161–179 (2014).
13) Morita S, Hourai A, Miyata S. Changes in pericytic expression of NG2 and PDGFRB and vascular permeability in the sensory circumventricular organs of adult mouse by osmotic stimulation. Cell Biochem Funct, 32(1), 51–61 (2014).
14) Morita S, Ukai S, Miyata S. VEGF-dependent continuous angiogenesis in the median eminence of adult mice. Eur J Neurosci, 37(4), 508–518 (2013).
15) Nishikawa K, Furube E, Morita S, Horii-Hayashi N, Nishi M, Miyata S. Structural reconstruction of the perivascular space in the adult mouse neurohypophysis during an osmotic stimulation. J Neuroendocrinol, 29(2), (2017).
16) Morita-Takemura S, Nakahara K, Tatsumi K, Okuda H, Tanaka T, Isonishi A, Wanaka A. Changes in endothelial cell proliferation and vascular permeability after systemic lipopolysaccharide administration in the subfornical organ. J Neuroimmunol, 298, 132–137 (2016).
17) Langlet F, Levin BE, Luquet S, Mazzone M, Messina A, Dunn-Meynell AA, Balland E, Lacombe A, Mazur D, Carmeliet P, Bouret SG, Prevot V, Dehouck B. Tanycytic VEGF-A boosts blood-hypothalamus barrier plasticity and access of metabolic signals to the arcuate nucleus in response to fasting. Cell Metab, 17(4), 607–617 (2013).
18) Miyata S, Morita S. A new method for visualization of endothelial cells and extravascular leakage in adult mouse brain using fluorescein isothiocyanate. J Neurosci Methods, 202(1), 9–16 (2011).
19) Morita S, Miyata S. Different vascular permeability between the sensory and secretory circumventricular organs of adult mouse brain. Cell Tissue Res, 349(2), 589–603 (2012).
20) Morita S, Furube E, Mannari T, Okuda H, Tatsumi K, Wanaka A, Miyata S. Heterogeneous vascular permeability and alternative diffusion barrier in sensory circumventricular organs of adult mouse brain. Cell Tissue Res, 363(2), 497–511 (2016).
21) Willis CL, Garwood CJ, Ray DE. A size selective vascular barrier in the rat area postrema formed by perivascular macrophages and the extracellular matrix. Neuroscience, 150(2), 498–509 (2007).
22) Faraci FM, Choi J, Baumbach GL, Mayhan WG, Heistad DD. Microcirculation of the area postrema. Permeability and vascular responses. Circ Res, 65(2), 417–425 (1989).
23) Morita S, Miyata S. Accessibility of low-molecular-mass molecules to the median eminence and arcuate hypothalamic nucleus of adult mouse. Cell Biochem Funct, 31(8), 668–677 (2013).
24) Schaeffer M, Langlet F, Lafont C, Molino F, Hodson DJ, Roux T, Lamarque L, Verdié P, Bourrier E, Dehouck B, Banères JL, Martinez J, Méry PF, Marie J, Trinquet E, Fehrentz JA, Prévot V, Mollard P. Rapid sensing of circulating ghrelin by hypothalamic appetite-modifying neurons. Proc Natl Acad Sci USA, 110(4), 1512–1517 (2013).
25) Morita-Takemura S, Nakahara K, Hasegawa-Ishii S, Isonishi A, Tatsumi K, Okuda H, Tanaka T, Kitabatake M, Ito T, Wanaka A. Responses of perivascular macrophages to circulating lipopolysaccharides in the subfornical organ with special reference to endotoxin tolerance. J Neuroinflammation, 16(1), 39 (2019).
26) Langlet F, Mullier A, Bouret SG, Prevot V, Dehouck B. Tanycyte-like cells form a blood-cerebrospinal fluid barrier in the circumventricular organs of the mouse brain. J Comp Neurol, 521(15), 3389–3405 (2013).
27) Morita-Takemura S, Wanaka A. Blood-to-brain communication in the hypothalamus for energy intake regulation. Neurochem Int, 128, 135–142 (2019).

This page was created on 2020-10-28T11:28:56.742+09:00
This page was last modified on 2020-12-21T10:12:42.000+09:00
このサイトは(株)国際文献社によって運用されています。