背側線条体におけるアストロサイト–ニューロン連関の機能解析
カリフォルニア大学ロサンゼルス校医学部生理学科
発行日:2020年12月30日Published: December 30, 2020
© 2020 日本神経化学会© 2020 The Japanese Society for Neurochemistry
生物が外界の情報を受容・処理し、次の行動に反映するしくみの理解は、脳科学の中心的な課題である。大脳皮質による認知的な随意運動と脳幹による生得的な運動は、大脳基底核により協調的に制御され、適切な運動機能の発現に寄与する。大脳基底核の主要な構成要素である線条体は、運動調節・認知・強化学習に関わる。またパーキンソン病・ハンチントン病・薬物依存などにおいて機能変調を示すことが知られている1–3)。このように個体行動や疾患に関与する線条体の機能は、ニューロン・グリア細胞・血管など多様な細胞の中、あるいは間で起こるダイナミックなシグナル伝達が統合されて発揮されると考えられる。しかし、細胞間相互作用のメカニズムおよび意義については不明な点が多い。
アストロサイトは中枢神経系にタイル状にひしめくグリアの一種である。哺乳類の脳細胞の約2~4割を占め、ニューロンと密なコミュニケーションをとっている4)。その機能に関して、大脳皮質や海馬での研究が数多く行われてきたが、近年の研究から背側線条体アストロサイトについての知見が集まっている。まず、アストロサイト微細突起はニューロンおよびシナプスと緊密に接している5, 6)。また、アストロサイト特異的RNAシーケンス(RNA-seq)により得られた遺伝子発現データからは、代謝補助、イオン・伝達物質のバッファリングといった、脳回路機能を支えるホメオスタティックなアストロサイト機能が示唆されている5)。さらに、アストロサイトでは電気的活動シグナルの伝搬が観察されないが、ダイナミックな細胞質Ca2+イオンの増減が見られる(Ca2+シグナル)ことが分かってきた7)。興味深いことに、アストロサイトのホメオスタティックな機能分子およびCa2+シグナルは、ハンチントン病モデルマウスで減少していることが明らかになっている7, 8)。
しかし、背側線条体アストロサイトの機能ダイナミクスについては、その多くが不明であった。特に、(1)アストロサイトがいつ・どのようにニューロン活動に呼応するのか、(2)その結果、どのような機能を発揮し、神経回路・個体行動に影響するのか、という根本的な生理学的メカニズム解析がなされていなかった。筆者は、分子・回路・動物行動レベルの多階層解析を通して、背側線条体におけるアストロサイト-ニューロンの相互作用のしくみと意義の解明を目的として研究を行った9, 10)。本稿では、まず(1)ニューロンからアストロサイトへのシグナル伝達機構、次に(2)アストロサイトのCa2+シグナルを「不活性化」「活性化」した際の行動・回路への影響について概説する。
これまで、ニューロン活動により放出される伝達物質(グルタミン酸、ATP、カンナビノイドなど)や修飾物質(ノルアドレナリン、アセチルコリンなど)がアストロサイトCa2+シグナルを上昇させることが報告されてきた11)。しかし、成体マウス背側線条体での機構は不明であった。そのため、まず成体マウス脳スライスを用いて、ニューロンの興奮性操作とアストロサイトCa2+シグナルイメージングを同時に行った(図1A)。ホールセルパッチクランプにより中型有棘ニューロン(MSN)をup-state様(静止膜電位より+20–30 mV)興奮12)を引き起こしたところ、近傍(細胞体か樹状突起から50 µm以内)に存在するアストロサイトがCa2+シグナル上昇を示した(図1B)。驚くべきことに、TTXで活動電位を阻害してもこのCa2+シグナルは上昇した。ニューロンの電位変化のみでアストロサイトCa2+シグナルを引き起こすメカニズムとして、樹状突起に高発現しているL-type電位依存性Ca2+からニューロンへCa2+が流入13)し、小胞放出を介して伝達物質がアストロサイトに作用していることを薬理学的実験により見出した。
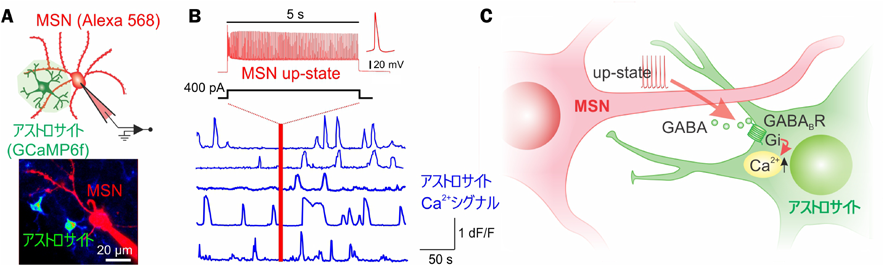
A, 赤色色素ダイアライシスによって可視化されたMSNとCa2+指示薬GCaMP6fを発現したアストロサイト。B, up-state様の興奮性を模倣したMSN脱分極の後、アストロサイトCa2+シグナル(青)の頻度が増加した。C, MSNの脱分極依存的なGABA放出は、アストロサイトが発現するGABAB受容体を介してGi経路を活性化し、Ca2+シグナルを引き起こす。
次に、MSNはGABA作動性ニューロンであることから、アストロサイトCa2+シグナルを上昇させる伝達物質はGABAではないかと仮説を立てた。生体内から単離したアストロサイトのmRNAおよびタンパク質の解析から、GABAB受容体が高発現していることが見出された。これを受けて、GABAおよびGABAB受容体アゴニストを急性スライスのアストロサイトに投与したところ、Ca2+シグナルを引き起こされた。また、MSN興奮性依存的なアストロサイトCa2+シグナルはGABAB受容体ノックダウンにより阻害された。さらに、光遺伝学的にin vivoニューロンを興奮させた場合においても、GABAB受容体を介したアストロサイトGi/Ca2+シグナルの上昇が観察された。これらのことから、MSNのup-state様活動依存的に放出されるGABAが、アストロサイトが高発現するGi共役型GABAB受容体を活性化させ、Ca2+シグナルを上昇させることが示された9)(図1C)。
では上記のアストロサイトCa2+シグナルはどのような機能的意義をもつのであろうか? この問いに答えるための実験的アプローチを考案する際に、以下のような留意点が存在した。まず、GABAB受容体はアストロサイトで高発現しているとはいえニューロンや脳以外の臓器にも発現があるため、例えばGABAB受容体アゴニストをマウスに投与して観察される表現型は、アストロサイトGABAB受容体刺激のみを反映した結果であるとみなすことは難しい。また、Creライン14)を用いてGABAB受容体をアストロサイト特異的に欠損させることも可能だが、欠損は背側線条体以外の多領域に及ぶため、例えば欠損マウスで行動変化が見られた場合に、背側線条体依存的である部分を切り分けることが困難である。脳領域特異的なアストロサイトマーカー/プロモーターの発見は今後の重要な課題である15)。
以上のことから、アストロサイト特異性・脳領域選択性を兼ね備えたGfaABC1D指向性アデノ随伴ウィルス(AAV)を用いたアプローチを採用した9)(図2)。アストロサイト特異的かつ背側線条体選択的に、人工のGi共役型GPCRであるGi-DREADD hM4Di16)を発現させ、これを刺激する合成リガンドの投与により、時空間的に制御した形で1で示したシグナルを模倣する系を確立した(図2A)。
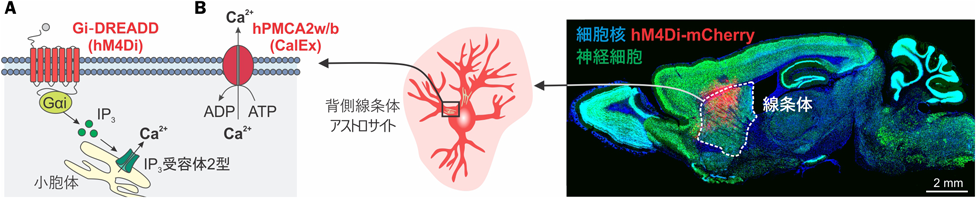
背側線条体アストロサイトにGi-DREADD刺激を与えたマウス(以下hM4Diマウス)では、合成リガンド投与2時間後にアストロサイトCa2+シグナルが上昇することが示された。また、hM4Diマウスはオープンフィールド試験において、落ち着きなく動き続ける様子(多動)が観察された。線条体が担う機能の一つである運動調節を調べるために、Rotarod試験を行ったところ、運動記憶には問題が見られなかったいっぽう、hM4Diマウスは落ち着きのない行動のためrotarod上から落下することが散見された。これを受け、注意欠陥を調べる目的で、視覚的な刺激(光、新奇物体)に依存する反応性を検証したところ、hM4Diマウスは顕著に鈍い反応性を示した。計10種の行動試験の結果を統合して、この行動表現型は注意欠陥・多動性障害(ADHD: Attention deficit/Hyperactivity disorder)を想起させる表現型であると結論づけた。
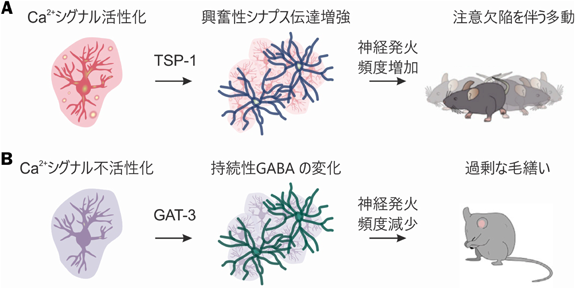
さらに、アストロサイトGi-GPCR刺激がADHD様行動を引き起こすメカニズムを解明した。アストロサイト特異的RNA-seq、脳スライス電気生理学、覚醒マウスにおけるニューロン活動記録を組み合わせた結果、ADHD様行動を示しているマウスでは、アストロサイトがシナプス産生因子TSP-1を放出し、線条体における皮質–線条体経路の興奮性シナプス過剰産生を引き起こしていることが見出された。TSP-1は神経系発生期に高発現し、回路形成に寄与する分子であり、成体マウスでは発現が非常に低いことが知られていた17)。しかし、アストロサイトGiシグナルの活性化はTSP-1を“再活性化”し、皮質–線条体経路の興奮性シナプス伝達を増強し、神経回路活動を亢進させることが明らかになった。TSP-1のニューロン受容体α2δ-1拮抗薬であるガバペンチン(ガバペン®)をマウスに投与したところ、異常なシナプス産生、回路活動亢進および行動が全て正常化された。以上の結果により、アストロサイトGiシグナルの活性化は発生期シナプス産生因子TSP-1を再活性化し、神経回路・個体行動に変化をもたらすことが見出された9)(図3A)。
以上の実験では、アストロサイト活性化による影響を検証したが、アストロサイトCa2+シグナルを減少させるアプローチでもその意義に迫った。アストロサイトCa2+シグナルの減少が確認されているマウスとしてIP3受容体2型欠損マウスが知られているが、このマウスの表現型については長らく議論がある18, 19)ことに加え、背側線条体特異的な欠損ではない。脳領域選択性を担保するために、AAVにより細胞膜上で機能するCa2+ポンプ(hPMCA2w/b)を過剰発現させ、Ca2+シグナルを低減させる新規の手法をCalEx(Calcium extruder)と名付けた(図2B)。AAVによる背側線条体アストロサイト特異的CalEx発現は、アストロサイト自発的Ca2+シグナルおよびGPCR依存的Ca2+シグナルを70–80%低減させることに成功した。
AAV注入の3週間後にCalExマウスの行動を複数の試験により解析したところ、過剰な毛繕い行動を示すことが見出された。単位時間当たりの毛繕い回数に変化はなく、毛繕いの時間が顕著に長くなっていた。高架式十字迷路試験の結果からは不安様行動が観察されなかったため、自らの意思に反して繰り返しの行動をしてしまう強迫性障害(OCD: Obsessive–compulsive disorder)を想起させる行動変化であると結論付けた。
この行動変化を引き起こすメカニズムを解明するために、脳スライス電気生理学を用いて、CalExマウスにおけるMSNのシナプス伝達記録を行った。その結果、興奮性伝達には影響がなく、持続性抑制が低減されていることが見出された。このシナプス表現型はアストロサイトGABAトランスポーターGAT-3の阻害剤により消失した。興味深いことに、CalExによるOCD様行動、またアストロサイトCa2+シグナル低減を伴うハンチントン病モデルマウスにおけるOCD様行動もGAT-3阻害剤によって有意に抑制されることが分かった。CalExマウスのアストロサイトRNA-seq解析から、背側線条体アストロサイトの自発的Ca2+シグナルは、神経伝達物質バッファリングを含むホメオスタティックな機能を維持するための遺伝子発現を調節しており、その変調は精神障害様の行動変化を引き起こす可能性が見出された(図3B)。
上記の一連の研究を通して、成体マウスの背側線条体におけるアストロサイトの活性化と不活性化がどちらも精神疾患様の行動異常を引き起こすことが明らかになった。さらに、薬理学的に標的可能な細胞間相互作用メカニズムを見出すことができた。特に、上述のガバペンチンはすでに抗てんかん薬として使用されているため、今回の発見はドラッグリポジショニングによる新規治療戦略に貢献できる可能性がある。今後は、操作時間の分解能やシグナル経路特異性が改良されたツールなどの開発を試み、それらを用いて異なる脳領域においてアストロサイトに摂動を与えることで、アストロサイトの行動神経学的役割と意義が紐解かれていくことが期待される。グリア生理学の発展に伴い、将来的にニューロンに着目するだけでは見えてこなかった脳回路機能のしくみや疾患治療・診断・予防法の理解に寄与することが展望される。
本稿で紹介いたしました研究成果は、カリフォルニア大学ロサンゼルス校(UCLA)医学部生理学科で得られました。多大なるご指導を賜りましたBaljit Khakh教授と研究室内でのコラボレーション、ディスカッションをしていただいたKhakh研究室メンバーに厚く御礼申し上げます。また、共同研究先のMichael Fanselow教授、Sotiris Masmanidis准教授、Giovanni Coppola教授(全てUCLA所属)に、それぞれ動物行動学、In vivo電気生理学、RNA-seq解析を学ばせていただいたことに心より感謝申し上げます。最後に、本稿執筆の機会を与えて下さいました日本神経化学会選考委員の先生方関係者の方々、並びに関係者の先生方に深く感謝申し上げます。
1) Graybiel AM, Grafton ST. The striatum: Where skills and habits meet. Cold Spring Harb Perspect Biol, 7(8), a021691 (2015). doi: 10.1101/cshperspect.a021691
2) Kreitzer AC, Malenka RC. Striatal plasticity and basal ganglia circuit function. Neuron, 60(4), 543–554 (2008). doi: 10.1016/j.neuron.2008.11.005
3) Zhai S, Tanimura A, Graves SM, Shen W, Surmeier DJ. Striatal synapses, circuits, and Parkinson’s disease. Curr Opin Neurobiol, 48, 9–16 (2018). doi: 10.1016/j.conb.2017.08.004
4) Ventura R, Harris KM. Three-dimensional relationships between hippocampal synapses and astrocytes. J Neurosci, 19(16), 6897–6906 (1999).
5) Chai H, Diaz-Castro B, Shigetomi E, Monte E, Octeau JC, Yu X, Cohn W, Rajendran PS, Vondriska TM, Whitelegge JP, Coppola G, Khakh BS. Neural circuit-specialized astrocytes: Transcriptomic, proteomic, morphological, and functional evidence. Neuron, 95(3), 531–549 (2017). doi: 10.1016/j.neuron.2017.06.029
6) Octeau JC, Chai H, Jiang R, Bonanno SL, Martin KC, Khakh BS. An optical neuron–astrocyte proximity assay at synaptic distance scales. Neuron, 98(1), 49–66.e9 (2018). doi: 10.1016/j.neuron.2018.03.003
7) Jiang R, Diaz-Castro B, Looger LL, Khakh BS. Dysfunctional calcium and glutamate signaling in striatal astrocytes from huntington’s disease model mice. J Neurosci, 36(12), 3453–3470 (2016). doi: 10.1523/JNEUROSCI.3693-15.2016
8) Diaz-Castro B, Gangwani MR, Yu X, Coppola G, Khakh BS. Astrocyte molecular signatures in Huntington’s disease. Sci Transl Med, 11(514), eaaw8546 (2019). doi: 10.1126/scitranslmed.aaw8546
9) Nagai J, Rajbhandari AK, Gangwani MR, Hachisuka A, Coppola G, Masmanidis SC, Fanselow MS, Khakh BS. Hyperactivity with disrupted attention by activation of an astrocyte synaptogenic cue. Cell, 177(5), 1280–1292.e20 (2019). doi: 10.1016/j.cell.2019.03.019
10) Yu X, Taylor AMW, Nagai J, Golshani P, Evans CJ, Coppola G, Khakh BS. Reducing astrocyte calcium signaling in vivo alters striatal microcircuits and causes repetitive behavior. Neuron, 99(6), 1170–1187.e9 (2018). doi: 10.1016/j.neuron.2018.08.015
11) Guerra-Gomes S, Sousa N, Pinto L, Oliveira JF. Functional roles of astrocyte calcium elevations: From synapses to behavior. Front Cell Neurosci, 11, 427 (2017). doi: 10.3389/fncel.2017.00427
12) Wilson CJ, Kawaguchi Y. The origins of two-state spontaneous membrane potential fluctuations of neostriatal spiny neurons. J Neurosci, 16(7), 2397–2410 (1996).
13) Carter AG, Sabatini BL. State-dependent calcium signaling in dendritic spines of striatal medium spiny neurons. Neuron, 44(3), 483–493 (2004). doi: 10.1016/j.neuron.2004.10.013
14) Srinivasan R, Lu TY, Chai H, Xu J, Huang BS, Golshani P, Coppola G, Khakh BS. New transgenic mouse lines for selectively targeting astrocytes and studying calcium signals in astrocyte processes in situ and in vivo. Neuron, 92(6), 1181–1195 (2016). doi: 10.1016/j.neuron.2016.11.030
15) Yu X, Nagai J, Khakh BS. Improved tools to study astrocytes. Nat Rev Neurosci, 21(3), 121–138 (2020). doi: 10.1038/s41583-020-0264-8
16) Roth BL. DREADDs for neuroscientists. Neuron, 89(4), 683–694 (2016). doi: 10.1016/j.neuron.2016.01.040
17) Allen NJ, Eroglu C. Cell biology of astrocyte-synapse interactions. Neuron, 96(3), 697–708 (2017). doi: 10.1016/j.neuron.2017.09.056
18) Fiacco TA, McCarthy KD. Multiple lines of evidence indicate that gliotransmission does not occur under physiological conditions. J Neurosci, 38(1), 3–13 (2018). doi: 10.1523/JNEUROSCI.0016-17.2017
19) Savtchouk I, Volterra A. Gliotransmission: Beyond black-and-white. J Neurosci, 38(1), 14–25 (2018). doi: 10.1523/JNEUROSCI.0017-17.2017

This page was created on 2020-10-28T14:53:18.804+09:00
This page was last modified on 2020-12-18T17:39:28.000+09:00
このサイトは(株)国際文献社によって運用されています。