C9orf72リピート伸長変異による前頭側頭葉変性症の分子病態
大阪大学大学院医学系研究科精神医学
発行日:2021年12月30日Published: December 30, 2021
© 2021 日本神経化学会© 2021 The Japanese Society for Neurochemistry
この度、2021年度日本神経化学会優秀賞にご選出いただきまして誠にありがとうございました。選考委員長の等誠司先生より「神経化学」誌上に研究を紹介する機会をいただきました。またとない機会ですので会員の皆様に我々の研究を御紹介させていただきたいと思います。
はじめに少し自己紹介をさせていただきますと、私はもともと精神医学を志望して地元の愛媛大学医学部に入学したのですが、2回生のとき愛媛大学医学部細胞生理学(当時生理学第一)の田中潤也教授のもとでマイクログリアの神経化学的研究に参加させて頂いたのがきっかけで研究をはじめることになりました。最初の論文はラットの初代培養マイクログリアに機能的なノルアドレナリンの受容体が発現していて、LPS刺激によるマイクログリア活性化をノルアドレナリンが抑制するという趣旨のものでした1)。当時、慶應義塾大学の大学院生として藤田保健衛生大学におられた田中謙二先生(現 慶應義塾大学医学部先端医科学研究所教授)にお声掛けいただき、後に田中謙二先生が生理研に移られた際には、生理研のセミナーに参加させていただいて食事に連れて行っていただいたのも良い思い出です。
当時は全く予想していなかったことですが、後にこの論文はアルツハイマー病やパーキンソン病における青斑核ノルアドレナリン作動性ニューロンの障害が神経炎症を増悪させる機序との関連で注目を集め、現在まで多数引用していただいています。
さて私が現在研究している前頭側頭葉変性症(FTLD: frontotemporal lobar degeneration)は、前頭葉、側頭葉を中心とした神経変性により、緩徐進行性に脱抑制的な行動障害、人格の変化、言語の障害を来たす疾患です。臨床的には前頭側頭型認知症(FTD: frontotemporal dementia)と称され、FTLDは神経病理学的用語として用いられることが多いです。若年性認知症ではアルツハイマー型認知症に続いて2番目に頻度が高いとされています。厚生労働省の指定難病としても登録されており、病態メカニズムの解明、新たな診断方法、治療法、ケア方法の開発が求められています。
神経変性疾患の多くは、神経病理学的には蓄積タンパク質の種類や蓄積のパターンにより分類されますが、タウタンパク質を封入体の主成分とするFTLDはFTLD-Tau、またRNA結合タンパク質であるTDP-43を封入体の主成分とするFTLDはFTLD-TDPと称されます。
多くの神経変性疾患と同様に孤発性のものと家族性のものとが知られています。特に家族性のものについてはこの15年ほどの間に原因遺伝子が次々と見いだされてきました。
9番染色体と関連する遺伝型のFTLD家系が存在すること自体は以前から報告されていました。原因遺伝子変異は未同定でしたが、私がドイツ、ミュンヘン大学のChristian Haass教授の研究室に留学する前年の2010年には、疾患関連領域は既に3つの遺伝子のみを含む狭い領域にまでしぼられていました。ドイツ留学フェローシップをいただいたアレクサンダー・フォン・フンボルト財団への申請書のテーマは、この3つの遺伝子の機能解析をするというものでした。翌2011年に私が渡独する直前になって、この3つの遺伝子のうちの一つ、C9orf72(以下C9と略す)遺伝子という未知の遺伝子の、しかもイントロン領域に病原性遺伝子変異が同定されました。それはGGGGCCという6塩基モチーフの数百以上にも及ぶ繰り返し配列でした。そのため自然な形でこのリピート伸長変異の研究に取り組むこととなりました。Haass教授のラボでC9変異の解析に取り組むのは私一人しかおらず、ラボ内のRNA構造の専門家などと相談しながら研究を進めていくことになりました。
この変異ではFTDや筋萎縮性側索硬化症などの運動ニューロン疾患を発症します。同一家系内でもそれぞれを単独で発症したり、両者を合併したりします。またC9変異を有する患者やその血縁者には、前駆期に統合失調症や双極性障害などと診断されるような症例が高頻度にみられ、精神医学的にも興味深い遺伝子変異と思われます。
C9変異例の剖検脳では神経細胞の細胞質を中心にリン酸化をうけたTDP-43タンパク質の異常凝集が見られるため、神経病理学的には大きくFTLD-TDPに分類されました。そこで6塩基リピートがどのようなメカニズムでTDP-43の異常を引き起こすかについての研究がはじまりました。
他のイントロンリピート病からの類推から、リピートによって1)C9遺伝子の翻訳産物(C9ORF72タンパク質)の機能喪失が生じること、2)C9リピート転写産物(リピートRNA)がRNA結合タンパクを選択的に吸着隔離(sequestrate)し、その生理的機能を失わせること、という2つの病態機序が想定されました。後者に対応する所見として、剖検脳や患者由来の培養細胞では、リピートRNAとRNA結合タンパク質の共凝集物であるRNA fociと呼ばれる構造物がみられます。さらにC9変異例ではTDP-43の封入体とは別に、オートファジーのアダプターであるp62やユビキチンに対する抗体で陽性に染め出される一方で、TDP-43やαシヌクレインなどの既知の神経細胞内封入体構成成分に対する抗体では染め出されない謎の封入体が多数存在することも報告されました。
C9リピート変異の発見と同じ2011年にフロリダ大学のRanumらにより、もう一つ重要な論文がPNAS誌に報告されました。DNAはRNAへと転写され、さらにそのRNAがタンパクへと翻訳されます。翻訳には、「ここから翻訳を開始してください」というシグナルが必要であり、通常はそれがKozakルールに合致する周辺配列をともなった開始コドンAU(T)Gにあたります。Ranumらは一部のトリプレットリピート病において、本来の翻訳フレーム以外からの翻訳が開始コドン非依存性に生じているとするRepeat associated non-AUG translation(RAN翻訳)の存在を報告したのです。
この論文は当時、少なくとも私の周囲の認知症研究者の間ではさほど注目されておらず、論文の存在を伝えた際には、「過剰発現のアーチファクトをみているだけではないか」と皆否定的でした。しかし共同研究者であったDieter Edbauer教授とそのポスドクWeng Shih-Ming博士、そして私は、C9変異例にみられる謎の封入体が、6塩基リピートRNAが開始コドン非依存性に翻訳されて生じるリピートタンパク質に由来するのではないかと考え、研究を進めていくことにしました。ただ私自身もこのプロジェクトがうまくいく確率はそれほど高くないだろうと考えており、あくまでサイドプロジェクトとして研究を進めていきました。
最初にGGGGCCという6塩基のリピートが開始コドン非依存性の翻訳を受けた際に、どのようなタンパク質が生じるのかを検討したところ、GGG GCCフレームからはグリシン-アラニン(Gly-Ala)のリピートタンパク質が、1塩基ずれて、GGG CCGフレームからはグリシン-プロリン(Gly-Pro)のリピートタンパク質が、もう1塩基ずれて、GGC CGGフレームからはグリシン-アルギニン(Gly-Arg)のリピートタンパク質が産生されることが想定されました(図1)。そのため我々はそれぞれのリピートタンパク質に対する抗体を作成しました。私がそれらの抗体をミュンヘン大学の神経病理学教室に持ち込んで、同教室のThomas Arzberger博士が収集されているC9変異患者の剖検脳の海馬切片を染色させていただいたところ、非常にクリアなバックグラウンドに特徴的な形態の封入体のみが鮮やかに染め出されました。このときの感動は忘れられません。C9 FTLD/ALSの病原性の本態とも考えられるジペプチドリピートタンパク質(DPR: dipeptide repeat proteins)が姿を現した瞬間でした。

続いて患者由来のリピートを培養細胞に発現させると開始コドン非依存性にDPRが産生されること、患者小脳ライセートにDPRが含まれること、DPR封入体はTDP-43封入体とは別の封入体であることなどを示し、Science誌に投稿したところ、いくつかの改定実験を要求された後に正式に受理されました2)。さらにフォローアップの仕事としてGGGGCCリピートがアンチセンス方向、すなわちCCCCGGリピートとして転写、翻訳され、プロリン-アラニン(PA)リピート、プロリン-アルギニン(PR)リピートが生じることも示しました(図1)3)。
現在までにGAリピートおよびGR, PRリピートが神経毒性を有することが複数のグループから実験的に示されています(多数の論文があるのでここでは私が関与したもののみを示させていただきます4, 5))。特にGR, PRリピートというアルギニン残基を含むDPRは、細胞内の分子間のゆるやかな相互作用を介した相分離現象に強い影響を与えて毒性を発揮しています。リピートRNAもしくはDPRにより細胞質–核の間の輸送が障害されることも報告され、TDP-43の異常蓄積に繋がる病態メカニズムとして注目されています。
留学当初の私のメインプロジェクトはC9のGGGGCCリピートRNAに選択的に結合するタンパク質を同定し、RNA結合タンパク質の観点からC9 FTLD/ALSの病態に切り込むことでした。そのためにまずin vitro RNA結合アッセイを構築して、GGGGCCリピートRNAに選択的に結合するRNA結合タンパク質をHEK293細胞の核ライセートから生化学的にアフィニティ精製し、質量分析によりそれら一群のタンパク質を同定しました。得られた20種類のリピートRNA結合タンパク質のうち、市販抗体が利用できるものについて、患者脳切片を用いた免疫組織学的スクリーニングを行いました。するとhnRNPA3の抗体で染色した健常組織では細胞核が染色されるのに対し、C9患者組織では細胞核への染色性が一部失われ、さらに海馬歯状回に限局した封入体病理も観察されました6)。
続いて我々は、このhnRNPA3がC9 FTLD/ALS病態で果たす役割について多角的に検討しました。まず外来性にGGGGCCリピートを発現させた細胞においてhnRNPA3をノックダウンするとGGGGCCリピートRNAの蓄積レベルが増大することがわかりました。逆にhnRNPA3を過剰発現するとリピートRNAの発現レベルは低下しました。つまりhnRNPA3はリピートRNAの発現を抑制的に制御していることが明らかになりました。同様の知見が、患者由来の線維芽細胞を用いた解析やラット初代培養ニューロンを用いた解析、さらには患者組織の解析でも確認されました。hnRNPA3はおそらくリピートRNAの代謝を促進することでリピートRNAの発現レベルを抑制していること、hnRNPA3が失われこの抑制が外れるとリピートRNAが著しく蓄積し、DPRの発現レベルも増加するという仕組みが存在することを明らかにしました(図2)7, 8)。
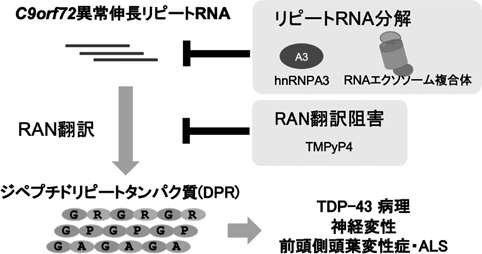
C9変異保持者由来の細胞では、転写されたC9orf72トランスクリプトの発現レベルは、変異をもたないものの約半分程度にとどまります。一方で同じトランスクリプトに由来するリピートRNAは、上述したRNA fociと呼ばれる構造物を形成して細胞内に蓄積しています。転写レベルが低下しているにもかかわらず、リピートRNAの蓄積がみられるという相反した現象から、我々はC9変異において異常伸長リピートRNAの分解が障害されている可能性を考えました。
こうした異常伸長リピートRNAが細胞内でどのように分解されるのかについての知見は全くなかったため、我々はいくつかの代表的なRNA分解酵素系を構成する分子をノックダウンしてその際のDPRの発現レベルを検証しました。その結果RNAエクソソーム複合体、特に核小体に局在するサブユニットであるEXOSC10がリピートRNAの分解に重要な役割を果たしていることを見出しました9)。核小体は細胞内で分子間のゆるやかな相互作用を介した液—液相分離とよばれる現象により形成される細胞内小器官であり、主にリボソームの生合成を担っています。アルギニン含有DPRであるGR, PRを発現させると核小体に集族してribosomal(r)RNA産生を阻害することも知られており、我々はRNAエクソソーム複合体に興味を深め、さらに解析を進めることにしました。
患者由来線維芽細胞においてEXOSC10をノックダウンすると細胞内にリピートRNAが蓄積し、それにともなって線維芽細胞の核におけるRNA fociも増加しました。これはEXOSC10/RNAエクソソーム複合体が患者細胞のリピートRNAを分解する酵素であることを意味します(図2)。リピートRNAを発現した細胞のうち、RAN翻訳によりアルギニン含有DPR(すなわちGR, PR)を発現した細胞では、EXOSC10が核小体に限局せず核内にびまん性に広がりました。またプルダウン実験により、EXOSC10はアルギニン含有GR, PRと相互作用をすることも示しました。
EXOSC10は、一部のsmall nucleolar(sno)RNA前駆体のプロセシングに関わっています。EXOSC10の基質として知られているsnoRNA48前駆体、およびsnoRNA68前駆体の蓄積レベルを評価したところ、GR, PR発現細胞では、コントロール細胞に比べて両前駆体ともにより多く蓄積していました。アルギニン含有DPRであるGR, PRが内因性のEXOSC10の活性を阻害していることを示していると考えられました。さらにこれらの細胞にGGGGCCリピートRNAを発現させたところ、上述したsnoRNA48前駆体やsnoRNA68前駆体と同様に、GGGGCCリピートRNAの蓄積が確認できました。
これらの結果から、アルギニン含有DPRであるGR, PRがrRNAの生合成の低下を引き起こすのみならず、EXOSC10/RNAエクソソーム複合体の活性低下を介して、リピートRNAの分解を阻害することを明らかにしました。リピートRNAの分解が阻害されることで、DPRの産生が加速し、病態を更に悪化させるものと想定されます9)。
DPRは強い細胞毒性を持つことが報告されているため、RAN翻訳を選択的に阻害し、DPRの産生を阻害することで新たな治療方法を開発できるかもしれません。私達はこのような観点からの研究にも取り組んできました。
GGGGCCリピートRNAはカリウムイオンの存在下でG-quadruplex(グアニン四量体)と呼ばれる強固な3次構造を取ることが知られています(図3)。TMPyP4(5,10,15,20-Tetrakis-(N-methyl-4-pyridyl)porphine)という化合物はポルフィリンの一種で、G-quadruplexに選択的に結合することが知られており、GGGGCCリピートRNAにも結合することが報告されていました。そこで我々はこのTMPyP4がGGGGCCリピートのRAN翻訳に及ぼす影響について検討しました10)。リピートを発現させた培養細胞にTMPyP4を処理したところ、TMPyP4はRAN翻訳によるDPRの産生を阻害しました。このときAUG依存性のEGFPの翻訳や培養細胞の翻訳活性全体は阻害されていなかったため、TMPyP4はRAN翻訳を選択的に抑制していることがわかりました。
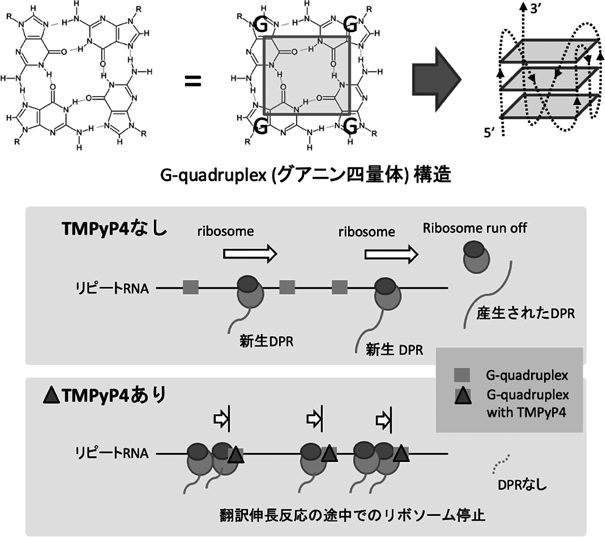
続いて、TMPyP4がRAN翻訳を選択的に抑制する機序を明らかにすることを試みましたが、TMPyP4はAUG非存在下、存在下いずれの場合でもリピートの翻訳を阻害したため、TMPyP4はRAN翻訳におけるAUG非依存性翻訳の開始を特異的に阻害しているというよりも、むしろRAN翻訳の伸長反応を阻害するのではないかと考えるに至りました。
翻訳の伸長反応が阻害された場合には、一つのRNA上に多数のリボソームが停止します。そのため細胞質をショ糖密度勾配法で分画した際には、比重の重い分画にRNAとリボソームの複合体が回収されます。TMPyP4で処理した細胞の細胞質をショ糖密度勾配法で分画してみると、予想通り未処理の細胞と比べて比重の重い分画にリピートRNAが多く局在することが明らかになりました。さらにTMPyP4とリピートRNAが変性尿素ゲルに耐性の強固な相互作用を示すことも明らかになりました。
これらの結果からTMPyP4はGGGGCCリピートRNAに強固に結合し、RAN翻訳の伸長反応を阻害することでRAN翻訳を阻害していることを示すことができました(図3)。将来的にはより選択性や結合の強さを高めたリピートRNA結合化合物によってRAN翻訳を阻害して、実際の治療に役立てることが可能になるのではないかと期待しています10)。
C9リピート伸長変異によるFTLD/ALSが見いだされてから今年でちょうど10年になります。この10年の間に分子病態に関して多くの知見が積み重ねられてきました。私も僅かながらそこに貢献できたのではないかと思いますが、これらの知見を患者さんやそのご家族に還元できるようになるまでには、さらなる研究が必要です。また近年次々と新規リピート病が見いだされてきており、リピート病の研究はさらなる発展が期待されます。私達の研究グループでは大学院生を募集していますので、興味のある方はぜひ私までご連絡をいただければと思います。
これまでに研究をご指導いただきました田中潤也先生、武田雅俊先生、大河内正康先生、田上真次先生、Christian Haass先生、Dieter Edbauer先生、Thomas Arzberger先生、永井義隆先生、現所属の大阪大学精神医学教室の池田学先生、そして研究室のメンバー達にこの場をお借りして深謝申し上げます。
1) Mori K, Ozaki E, Zhang B, Yang L, Yokoyama A, Takeda I, Maeda N, Sakanaka M, Tanaka J. Effects of norepinephrine on rat cultured microglial cells that express alpha1, alpha2, beta1 and beta2 adrenergic receptors. Neuropharmacology, 43(6), 1026–1034 (2002).
2) Mori K, Weng SM, Arzberger T, May S, Rentzsch K, Kremmer E, Schmid B, Kretzschmar HA, Cruts M, Van Broeckhoven C, Haass C, Edbauer D. The C9orf72 GGGGCC repeat is translated into aggregating dipeptide-repeat proteins in FTLD/ALS. Science, 339(6125), 1335–1338 (2013).
3) Mori K, Arzberger T, Grasser FA, Gijselinck I, May S, Rentzsch K, Weng SM, Schludi MH, van der Zee J, Cruts M, Van Broeckhoven C, Kremmer E, Kretzschmar HA, Haass C, Edbauer D. Bidirectional transcripts of the expanded C9orf72 hexanucleotide repeat are translated into aggregating dipeptide repeat proteins. Acta Neuropathol, 126(6), 881–893 (2013).
4) May S, Hornburg D, Schludi MH, Arzberger T, Rentzsch K, Schwenk BM, Grasser FA, Mori K, Kremmer E, Banzhaf-Strathmann J, Mann M, Meissner F, Edbauer D. C9orf72 FTLD/ALS-associated Gly-Ala dipeptide repeat proteins cause neuronal toxicity and Unc119 sequestration. Acta Neuropathol, 128(4), 485–503 (2014).
5) Zhou Q, Lehmer C, Michaelsen M, Mori K, Alterauge D, Baumjohann D, Schludi MH, Greiling J, Farny D, Flatley A, Feederle R, May S, Schreiber F, Arzberger T, Kuhm C, Klopstock T, Hermann A, Haass C, Edbauer D. Antibodies inhibit transmission and aggregation of C9orf72 poly-GA dipeptide repeat proteins. EMBO Mol Med, 9(5), 687–702 (2017).
6) Mori K, Lammich S, Mackenzie IR, Forne I, Zilow S, Kretzschmar H, Edbauer D, Janssens J, Kleinberger G, Cruts M, Herms J, Neumann M, Van Broeckhoven C, Arzberger T, Haass C. hnRNP A3 binds to GGGGCC repeats and is a constituent of p. 62-positive/TDP43-negative inclusions in the hippocampus of patients with C9orf72 mutations. Acta Neuropathol, 125(3), 413–423 (2013).
7) Mori K, Nihei Y, Arzberger T, Zhou Q, Mackenzie IR, Hermann A, Hanisch F, Kamp F, Nuscher B, Orozco D, Edbauer D, Haass CGerman Consortium for Frontotemporal Lobar DegenerationBavarian Brain Banking Alliance. German Consortium for Frontotemporal Lobar Degeneration, Bavarian Brain Banking Alliance, Kamp F, Nuscher B, Orozco D, Edbauer D, Haass C. Reduced hnRNPA3 increases C9orf72 repeat RNA levels and dipeptide-repeat protein deposition. EMBO Rep, 17(9), 1314–1325 (2016).
8) Nihei Y, Mori K, Werner G, Arzberger T, Zhou Q, Khosravi B, Japtok J, Hermann A, Sommacal A, Weber M, Kamp F, Nuscher B, Edbauer D, Haass C; German Consortium for Frontotemporal Lobar Degeneration; Bavarian Brain Banking Alliance. German Consortium for Frontotemporal Lobar Degeneration, Bavarian Brain Banking Alliance, Kamp F, Nuscher B, Edbauer D, Haass C. Poly-glycine-alanine exacerbates C9orf72 repeat expansion-mediated DNA damage via sequestration of phosphorylated ATM and loss of nuclear hnRNPA3. Acta Neuropathol, 139(1), 99–118 (2020).
9) Kawabe Y, Mori K, Yamashita T, Gotoh S, Ikeda M. The RNA exosome complex degrades expanded hexanucleotide repeat RNA in C9orf72 FTLD/ALS. EMBO J, 39(19), e102700 (2020).
10) Mori K, Gotoh S, Yamashita T, Uozumi R, Kawabe Y, Tagami S, Kamp F, Nuscher B, Edbauer D, Haass C, Nagai Y, Ikeda M. The porphyrin TMPyP4 inhibits elongation during the noncanonical translation of the FTLD/ALS-associated GGGGCC repeat in the C9orf72 gene. J Biol Chem, 297(4), 101120 (2021).

This page was created on 2021-11-24T11:23:09.836+09:00
This page was last modified on 2022-02-25T15:17:49.000+09:00
このサイトは(株)国際文献社によって運用されています。