精神疾患MRI研究のために神経化学研究者がなすべき義務について
慶應義塾大学 医学部先端医科学研究所 脳科学研究部門
発行日:2021年12月30日Published: December 30, 2021
© 2021 日本神経化学会© 2021 The Japanese Society for Neurochemistry
核磁気共鳴画像法(MRI)は非侵襲的に脳機能と脳構造の両方を全脳レベルで可視化出来る唯一無二の臨床機器である。精神疾患の臨床研究において、神経活動変化を可視化する機能的MRI(functional MRI: fMRI)、脳ネットワーク変化を可視化する安静時fMRI(resting state fMRI: rsfMRI)、脳微細構造変化を可視化する拡散MRI(diffusion MRI: dMRI)、脳体積変化を可視化するVBM(voxel-based morphometry)は必須の技術となっている。逆に、これらの技術を精神疾患の診断に用いられるかというと、答えはNoである。その理由として、MRI信号変化を脳科学的どのように解釈して良いのか曖昧であり、MRI信号変化と精神疾患の病態の関係性が曖昧だからである。
例えば、日本の精神科の共同研究チームは統合失調症患者では淡蒼球の体積が増加していることを発見した1)。しかし、統合失調症と淡蒼球体積増加の因果関係は全く分からない。また、脳体積の増加は脳科学的に見ると何が起こっているのか全く分からない。シナプスが増えたのか、神経細胞が増えたのか、またはグリア細胞が増えたのか、または細胞が大きくなったのか、MRIデータを見ただけでは全く分からない。rsfMRIにおいても同様なことが言える。ある2領域間のコネクションに変化があったとういうデータを得た場合、コネクションの変化は電気生理学的にどのように解釈できるのだろうか。コネクションが変化した脳領域と病態に関係性があるのだろうか。そもそもrsfMRIは血流変化を介したコネクションを見ているため、rsfMRIデータの結果をどのように解釈したら良いのだろうか。
このように、ただ単にMRIデータの結果を見ていただけでは、実際に脳では何が起こっているのか、その本質を理解することは極めて困難である。推測は可能だが、あくまでも推測である。しかし、日本だけではなく世界中で大規模な精神疾患を対象としたMRI研究が盛んに行われている。私は、一見デメリットしか見えないMRIをなぜこれだけ多用するのかと一時は疑問に思ったことがある。しかし、臨床研究にはMRIしかないという現実に気付された。「非侵襲性」「全脳探索性」「解析の多様性」というMRIのメリットは、上述したMRIのデメリットを圧倒していることに気付かされた。
そこで私は、MRI信号変化に脳科学的な解釈を与えること、そして病態とMRI信号変化に因果関係を持たせることの重要性について考えるようになった。これを達成するためには、病態を詳細に理解しなければならい。そして、その病態に介入し、MRI信号変化と因果関係があることを示さなければならない。病態を詳細に理解する為には、神経細胞やグリア細胞の形態を知る必要がある。そして、その形態を詳細に記述する観察技術が必要である。また、生理変化を理解するために、イメージング技術や電気生理学的な技術も必要である。血流についても知らなければならない。病態に介入する為には、遺伝子工学、薬理学、光遺伝学に精通しなければならない。これらの技術は全て神経化学研究者が得意とする技術であり、神経化学研究者が最前線に立って、取り組むべき課題、むしろ義務だと私は思っている。私の大きな研究目標は、神経化学を代表する技術を用いて、MRI信号変化に神経化学的な解釈を与えることである。そして、病態とMRI信号変化に因果関係を示すことである。将来的には、MRI信号変化を見ただけで、脳で何が起こっているのか理解できる基盤を構築したいと考えている。本稿では、fMRIに焦点を絞り、fMRIの弱点とそれを克服した新たなfMRI技術の紹介を行い、精神疾患とfMRI信号変化について議論したい。
fMRIはneurovascular couplingの原理を元に、血流動態変化を神経活動変化として捉える技術である(図1左)。しかし、血流動態に異常が生じると正確な神経活動を評価することが出来なくなってしまう2)。また、MRI技術の進歩により、画像の空間分解能が上昇することで、血管密度にfMRI信号(BOLD信号と呼ばれるが、本稿ではfMRI信号と呼ぶ)が依存することも分かってきた。さらに、神経活動マーカーであるc-fos遺伝子の発現分布とfMRI信号の分布が一致しないというのも有名である。fMRI信号が神経活動を表すということは周知の事実ではあるが、fMRI信号が血流変化を信号源とするため、fMRIによって得られたデータの解釈には複雑性が増してしまう。それでも、fMRIを用いた大規模臨床試験を行う重要性は極めて高く、fMRIデータを適切に解釈できるようにならなければならない。しかし、ここで私は全く逆の発想に至った。すなわち、血流を介さずにfMRIを遂行出来れば、この血流とfMRIの問題を解決できるのではないかと考えた。
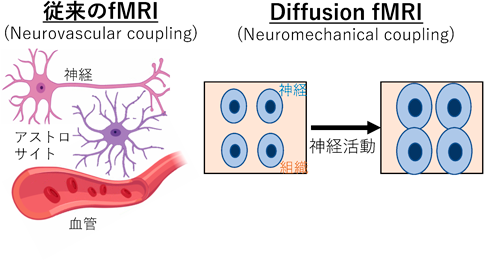
従来のfMRIは血流動態を介して神経活動を可視化する(neurovascular coupling)のに対して、DfMRIは細胞体積変化を介して神経活動を可視化する(neuromechanical coupling)。
私は神経活動が起こると細胞体積が僅かに増加することに着目した3, 4)。この僅かな体積増加の検出には、細胞浮腫や脳梗塞の検出として臨床研究で用いられるdMRIが最適である。dMRIは水の拡散の大きさを可視化する技術である5)。細胞体積が増加することで水の拡散の大きさも変化する。そこで神経活動を細胞体積変化としてdMRIによって捉えられるのではないかと考えた(図1右)。言い換えれば、神経活動を水の拡散で捉えられるのでないかということである。私は、この技術を既に考案していたdMRIの開発者の一人であるLe Bihan博士(NeuroSpin, フランス)の元に留学し、dMRIとfMRIを融合したdiffusion fMRI(DfMRI)という技術の確立を目指した。
Le Bihan博士のチームは既に、DfMRI信号(実際にはapparent diffusion coefficient: ADCというMRI信号を使用するが、本稿ではDfMRI信号と呼ぶ)が神経活動を反映する可能性があることを臨床試験によって示していた6)。また、DfMRI信号は血流非依存的であることも示していた7)。
はじめに、私はDfMRI信号が本当に神経活動を伴う細胞体積増加を捉えられるのかを検証するため、アメフラシの神経節を用いてMRIでの単一細胞イメージングに挑戦した8)。留学した研究施設(NeuroSpin, フランス)に設置されていた世界最高峰の超高磁場17.2 tesla MRIを用いた。まずはアメフラシを釣るところから実験がスタートする。ワインで有名なボルドー地方まで車を走らせ、浅瀬の海でアメフラシを採取した。アメフラシの神経節には直径200–500 µmの巨大な細胞体が存在する為、MRIでの単一細胞イメージングに適している(図2A)。神経節を培養及びかん流しながらMRI撮像可能なシステムを自作して実験を行った(図2B)。その結果、ドパミン刺激によって神経活動が増加した際に9)、DfMRI信号の減少を伴う体積増加を検出することができた(図2C–D)。今までの研究から、神経活動が増加するとDfMRI信号は減少することは既に分かっていた10)。この結果から、単一細胞イメージングにおいて、神経活動の増加によって細胞体積が増加することを示すことが出来た。そしてこの体積増加をDfMRIによって検出する事ができ、この時、DfMRI信号は減少することを示すことが出来た。
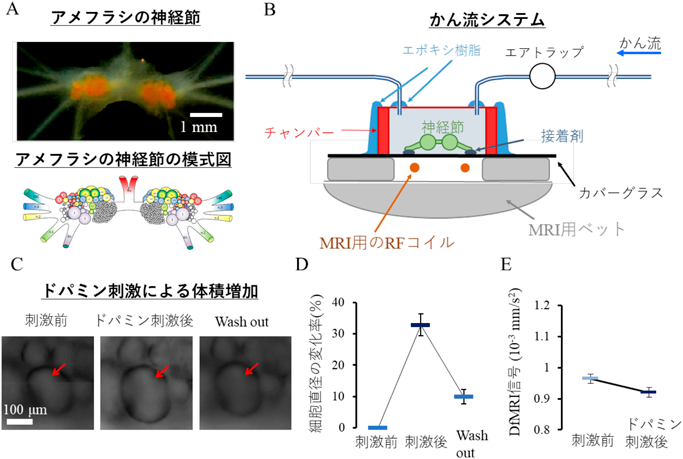
アメフラシの神経節(A)と、神経節を培養、かん流しながらのMRIシステム(B)。ドパミン刺激後の細胞体積変化を顕微鏡にて観察(C)。刺激による細胞直径変化(D)とDfMRI信号の変化(E)。
次に、神経活動を伴う体積増加を薬理学的に操作し、DfMRI信号が変化するのかを検証した2)。この実験では7 tesla MRIとラットを用いた。細胞体積増加を誘発させる低浸透圧人口脳脊髄液をラット脳内に局所投与し、経時的なDfMRI信号を観察した。また、局所投与した領域でlocal field potential(LFP)を計測した。その結果、神経活動の増加を伴うDfMRI信号の減少を検出することができた(図3上)。反対に、細胞体積を減少させる働きがあるfurosemide(NKCC1とKCC2のblocker)の局所投与によって、DfMRI信号を増加させることができた(図3下)。この結果から、DfMRI信号は神経活動を伴う体積増加を検出していることが分かった。
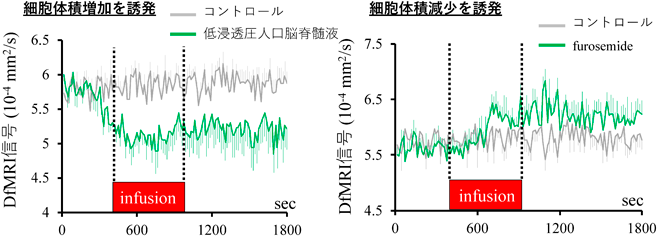
細胞体積増加を誘発する低浸透圧人口脳髄液を脳に局所注入し、DfMRI信号の減少を確認できた。細胞体積増減少を誘発するfurosemideを局所注入し、DfMRI信号の増加を確認できた。
最後に、血流動態に異常が起こっても、DfMRI信号は神経活動を正確に捉えられることを,同様の実験系を用いて確かめた2)。血流動態の異常は麻酔薬を用いた。イソフルランは血管拡張作用があり、血流を増加させる。麻酔濃度依存的なDfMRI信号の増加を検出できるのかを検証した。イソフルラン濃度依存的にLFPが減少することから、麻酔による神経活動の減少が起こっていることが分かる(図4上)。この時、血流増加に伴いfMRI信号は増加する。つまり、イソフルラン麻酔下(血管が拡張している状況)において従来のfMRIでは神経活動を適切に評価できないことが示された(図4下)。これに対して、DfMRI信号はイソフルラン濃度依存的に増加すること(つまり神経活動の減少を意味する)から(図4中)、血流動態に異常があっても神経活動を適切に評価できることを確認できた。
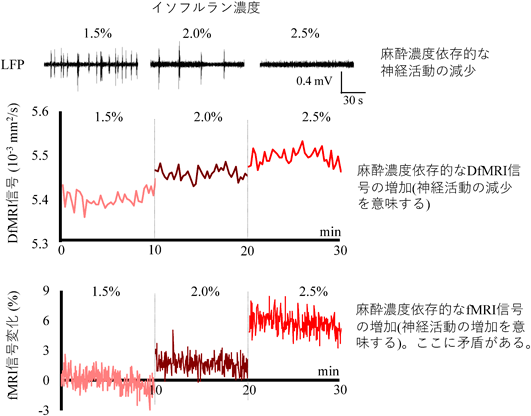
イソフルラン濃度依存的な神経活動変化(LFP)とDfMRI信号変化とfMRI信号変化の比較。
これらの実験から、DfMRI信号は1)血流動態に異常があっても神経活動を適切に評価できること、2)神経活動に伴う細胞体積変化を捉えられることを私は留学中に見いだすことができた。この技術を日本に持ち帰り、精神疾患モデルマウスへの応用を試みた。
繰り返しになるが、DfMRIの特徴をまとめる。一つ目は血流非依存的に神経活動を評価できることである。二つ目は神経活動を伴う細胞体積変化を検出していることである。DfMRIは上記2つの特徴に加え、もう1つ利点がある。fMRI信号は相対的な信号であり、ベースラインから信号がどれだけ変化したのか、その変化率を神経活動とするため、個体内の比較が主である。fMRI信号そのものの値を比較することは出来ない。これに対して、DfMRI信号であるADCという信号は、絶対値として扱うことができるため、DfMRI信号の大きさをそのまま比較することにより、個体間の比較が可能となる。つまり、DfMRI信号の大きさを安静時における神経活動(basal activity)の大きさとして、神経活動を評価することが出来る。これがDfMRIの特徴の三つ目である。これを用いることで、例えば、精神疾患モデルマウスのbasal activityの変化を可視化することが出来る。これは従来のfMRIでは不可能な技術である。
次に、DfMRIの特徴を生かし、血流非依存的なネットワーク解析が出来るのではないかと考えた。従来のfMRIでは、2領域間におけるfMRI信号の相関を見ることで機能的な結合(functional connectivity: FC)を評価することが出来る。DfMRIでも同様にFCも求め、ネットワーク解析が可能だと考えた。しかし、このままでは従来のfMRI研究と何も変わらない。神経活動変化とネットワークの変化を見出して終わりである。私はここからさらにもう一歩、何か踏み込みたかった。そこで、私は神経活動変化とコネクション変化に因果関係を持たせられないかと考え、神経活動が変化した瞬間、そこから伝搬される神経活動がどのように脳内に広がっていくのかを可視化することを目指した。この解析手法をignition-driven mean integration(IDMI)解析と呼ぶ。ある領域のある時刻において起こった神経活動がどれくらいの脳領域に広く伝搬するのかを可視化する解析である(図5)。理論は既に他のグループによって考案されており11)、私はこの理論をDfMRIに実装した。この理論の実装には沖縄科学技術大学院大学(OIST)の銅谷賢治教授と国際電気通信基礎技術研究所(ATR)の酒井雄希博士にご指導を頂いた。私はDfMRIの利点を生かし、血流非依存的な脳機能解析の以下の3つのパイプラインを構築した12)。①安静時におけるbasal activityの解析、②ネットワーク解析、③神経活動の伝搬解析。
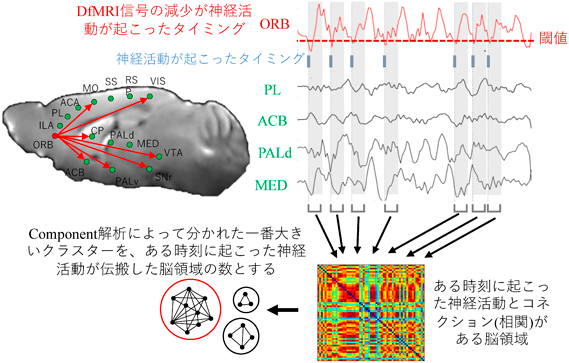
ある脳領域で、ある時刻に起こった神経活動がどれだけ多くの脳領域に、その神経活動が伝搬するのかを解析する。
アストロサイト特異的にグルタミン酸トランスポーターGLT1をノックダウンさせたマウス(GLASTCreERT2/+::GLT1flox/flox)は強迫症様症状(グルーミングやチック)が顕著に見られる13)。私はこのマウスを東京医科歯科大学の田中光一教授から供与を受け、解析した。このマウスはアストロサイトに起因したneurovascular couplingに異常が生じ、従来のfMRIでは正確な脳機能を評価できないことが想定されるため、neurovascular couplingに依存しないDfMRIの利点を最大限に生かすことができる。このモデルマウスを用いて上述したDfMRIの3つのパイプライン解析を行った。なお、MRIはOISTに導入されている11.7 tesla MRIを用いた。
その結果、眼窩前頭皮質(ORB)、前頭前野(PL/ILA)、運動野(MO)、島皮質(AI)、線条体(CP)、側坐核(ACB)、淡蒼球(PALd, PALv)、視床(MED)でbasal activityの増加(図6A)、cortico-striato-thalamo-cortical circuit(CSTC回路)のコネクション異常(図6B)、CSTC回路での神経活動伝搬の異常を発見することができた(図6C)。これらの異常はGLT1の減少の程度や強迫症様症状(グルーミング)の程度と相関も見られた。これに対して、通常のfMRIではこれらの異常を検出することが出来なかった。強迫症患者の臨床fMRI研究から、眼窩前頭皮質の過活動やCSTC回路の異常が報告されており、本研究のモデル動物を用いた結果と一貫性がある。以上のことから、この精神疾患モデルマウスにおいて、過活動、ネットワーク異常、活動伝搬の異常を可視化することに成功した。
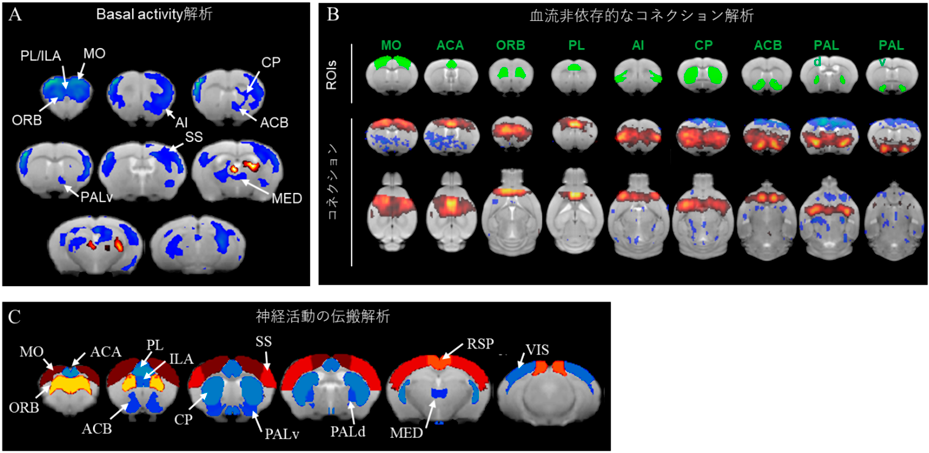
(A) DfMRI信号が減少した領域(basal activityが増加した領域)を青色で示す。(B)各脳領域にROI(緑)を置いた時、赤色(青色)はその領域と正(負)の相関がある領域を示す。(C)赤色はその領域から神経活動が伝搬した領域が多いことを示す。青色は少ないことを示す。
本稿ではfMRIに焦点を絞り、話を進めてきた。新規fMRIによって可視化出来た脳機能変化と病態を関連付けることに成功した。本稿で行ったDfMRI研究のように、DfMRI信号のメカニズムを理解する基盤研究から、そのDfMRI信号変化を病態と関連付ける応用研究まで展開出来れば、MRI研究の躍進に繋がると期待できる。
正直なところ、私は今のMRI研究がもったいないと感じている。現状では精神疾患患者を大量にリクルートし、VBMやrsfMRIのデータを集め、ある脳領域の体積やコネクションが変わったというデータを出している。しかし、それ以上の発見は何もない。MRI信号変化に神経化学的な解釈を与えることが出来れば、MRIデータから実際に脳で起こっていることを理解でき、精神疾患の病態理解に繋がる。さらにMRI研究の利点の一つとして、MRI装置は臨床と基礎で同じ装置、同じ技術を使用していることから、トランスレーショナル研究とリバーストランスレーショナル研究の双方向を行き来した研究が容易であり、精神疾患研究が加速度的に発展することが期待できる。将来的には、MRI信号変化から脳機能変化や脳構造変化を読み解けるようになることを期待している。欲を言えば、MRI信号変化から遺伝子変化まで読み解けるようになれば、申し分ないのだが、これは極めて難しいであろう。本稿で紹介した研究が少しでもその礎になれば幸いである。
この研究は上述した私の研究目標を達成する上での一例である。MRI研究はfMRIだけではなく、VBMやdMRIなど構造MRIもあり、これらのMRI信号変化も読み解く必要がある。構造MRIに関係する話は、優秀賞の時に熱く語れればと思っている。
日本神経化学会が掲げている「物質・分子・病態」と私が得意とするマウスMRI研究を融合させた研究で、このような評価を得られたこと、とても光栄に思っております。本稿で紹介しました研究成果は、NeuroSpin, Commissariat à l’énergie atomique et aux énergies alternatives, FranceのDenis Le Bihan博士の下と、慶應義塾大学医学部・先端医科学研究所・脳科学研究部門の田中謙二教授の下で行われました。多大なるご指導を賜りました。心より感謝申し上げます。また、謙二ラボの皆様や多数の共同研究者の皆様にもご指導を賜りました。心より感謝申し上げます。最後に、本稿執筆の機会を与えて下さいました日本神経化学会優秀賞・奨励賞の先生方、並びに関係者の先生方に深く感謝申し上げます。
1) Okada N, Yahata N, Koshiyama D, Morita K, Sawada K, Kanata S, Fujikawa S, Sugimoto N, Toriyama R, Masaoka M, Koike S, Araki T, Kano Y, Endo K, Yamasaki S, Ando S, Nishida A, Hiraiwa-Hasegawa M, Kasai K. Abnormal asymmetries in subcortical brain volume in early adolescents with subclinical psychotic experiences. Transl Psychiat, 8(1), 254 (2018).
2) Abe Y, Tsurugizawa T, Le Bihan D. Water diffusion closely reveals neural activity status in rat brain loci affected by anesthesia. PLoS Biol, 15(4), e2001494 (2017).
3) Buckley DL, Bui JD, Phillips MI, Zelles T, Inglis BA, Plant HD, Blackband SJ. The effect of ouabain on water diffusion in the rat hippocampal slice measured by high resolution NMR imaging. Magn Reson Med, 41(1), 137–142 (1999).
4) Pal I, Nyitrai G, Kardos J, Heja L. Neuronal and astroglial correlates underlying spatiotemporal intrinsic optical signal in the rat hippocampal slice. PLoS One, 8(3), e57694 (2013).
5) Le Bihan D. Diffusion MRI: what water tells us about the brain. EMBO Mol Med, 6(5), 569–573 (2014).
6) Le Bihan D, Urayama S, Aso T, Hanakawa T, Fukuyama H. Direct and fast detection of neuronal activation in the human brain with diffusion MRI. Proc Natl Acad Sci USA, 103(21), 8263–8268 (2016).
7) Tsurugizawa T, Ciobanu L, Le Bihan D. Water diffusion in brain cortex closely tracks underlying neuronal activity. Proc Natl Acad Sci USA, 110(28), 11636–11641 (2013).
8) Abe Y, Nguyen KV, Tsurugizawa T, Ciobanu L, Le Bihan D. Modulation of water diffusion by activation-induced neural cell swelling in Aplysia Californica. Sci Rep, 7(1), 6178 (2017).
9) Svehla P, Bédécarrats A, Jahn C, Nargeot R, Ciobanu L. Intracellular manganese enhanced MRI signals reflect the frequency of action potentials in Aplysia neurons. J Neurosci Methods, 295, 121–128 (2018).
10) Tsrugizawa T, Abe Y, Le Bihan D. Water apparent diffusion coefficient correlates with gamma oscillation of local field potentials in the rat brain nucleus accumbens following alcohol injection. J Cereb Blood Flow Metab, 37(9), 3193–3202 (2016).
11) Deco G, Kringelbach ML. Hierarchy of Information Processing in the Brain: A Novel ‘Intrinsic Ignition’ Framework. Neuron, 94(5), 961–968 (2017).
12) Abe Y, Takata N, Sakai Y, Hamada H, Hiraoka Y, Aida T, Tanaka K, Le Bihan D, Doya K, Tanaka KF. Diffusion functional MRI reveals global brain network functional abnormalities driven by targeted local activity in a neuropsychiatric disease mouse model. Neuroimage, 223, 117318–117318 (2020).
13) Aida T, Yoshida J, Nomura M, Tanimura A, Iino Y, Soma M, Bai N, Ito Y, Cui W, Aizawa H, Yanagisawa M, Nagai T, Takata N, Tanaka KF, Takayanagi R, Kano M, Götz M, Hirase H, Tanaka K. Astroglial glutamate transporter deficiency increases synaptic excitability and leads to pathological repetitive behaviors in mice. Neuropsychopharmacology, 40(7), 1569–1579 (2015).

This page was created on 2021-11-24T11:27:15.39+09:00
This page was last modified on 2022-02-25T15:25:48.000+09:00
このサイトは(株)国際文献社によって運用されています。