RNA結合蛋白質解析の利と美—柔軟かつ堅牢性の高い脳の組織、細胞機能を包括的に理解する—
新潟大学医学部神経解剖学
発行日:2022年6月30日Published: June 30, 2022
© 2022 日本神経化学会© 2022 The Japanese Society for Neurochemistry
近年のRNA結合蛋白質研究の歴史は、2012年のCastelloらの生化学的なmRNAキャプチャー法(後述するHITS-CLIPの逆のような方法)などで改めて同定、精査、分類され、2014年のGerstbergerらの総説にて、ヒトで1542種類と定義することが現在の定番となっている1, 2)。
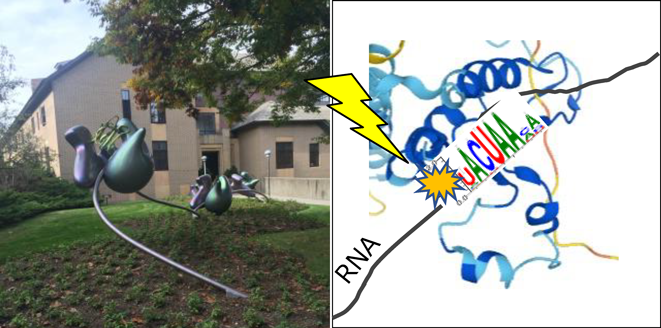
私とRNA結合蛋白質の関係は、本学会岡野栄之理事長との出会いにある。神経発生の種間を越えた統一的原理の抽出を夢見て大学院の研究をスタートしたわけだが、岡野栄之先生に提示されたこのRNA結合蛋白質の分子群を一瞬にして“超面白い”と直感した。ところが、この分子群は“分かった”と思うと、またすぐに仮説の更新を余儀なくされる複雑難解性を持ち合わせていた。まさに、大学院時代は暗闇から手を伸ばす思いで、神経特異的RNA結合蛋白質Elavlファミリー分子の機能解析に従事した。生化学、質量分析、ノックアウトマウスというツールを与えていただいて解析を進め、神経細胞の分化に関わるごくごく一部の機能を観察した3, 4)。ところがこの“分かった”はとても脆弱なもので、新たな“仮説の更新”と思考を巡らす必要があった。そんな中、幸運にも大学院時代のメンターであった岡野ジェイムス洋尚先生(現東京慈恵会医大教授)と共に研究成果を持ってコールドスプリングハーバーの学会発表をする機会をいただけた。ニューヨークの大都会、そして長閑な住宅街にあるコールドスプリングハーバー研究所の科学と一流科学者たちの匂いに興奮し、中でも私は、mRNAとリボソームの相互作用を介した蛋白質合成の瞬間をスナップショットするオブジェの美しさ(図1左)に目を奪われた。この時、岡野栄之先生からRNA結合蛋白質を提示された時の直感が確信に変わり、神経発生に重要なRNA結合蛋白質を見つけ出し、このオブジェのような解像度で細胞機能を全て手にとるように明らかにしたいという希望を強く持った。この夢の実現に向けて、私と家族まるごと包んでくれるようにサポートしてくれたのが、ロックフェラー大学ロバート・ダーネル博士(NYゲノムセンター創設者)であります。コールドスプリングハーバーに格好いい革ジャン姿にバイクで颯爽と現れて、このHuとhnRNP KはRNAのどことどこに結合してどれくらい離れているんだ?と聞かれたのを今でもよく覚えている。
本寄稿では、私たちがここ10年間で報告してきたRNA結合蛋白質研究とその解析技術から得られた成果について紹介したい。また、RNA結合蛋白質の機能とその破綻に伴う神経系の疾患と、RNA結合蛋白質解析の未来についても考えたい。
RNA結合蛋白質は、各々が膨大なRNA標的群を持ち、かつ多岐にわたるRNA制御機構を有することで、遺伝子発現制御の複雑性の増大に寄与している。それ故、RNA結合蛋白質はより柔軟な遺伝子制御を介し、細胞や組織にとって高度な堅牢性を担保する分子群であると言える。また、RNA結合蛋白質が様々な神経変性疾患や精神疾患の原因/病態関連分子であることから、遺伝子発現情報の柔軟性を失った細胞機能が破綻へと至る分子機構を理解するには、常に新たな技術革新と仮説の更新をし、精密かつ正確な包括的な分子ネットワークの解明が必須であることは言うまでもない。その中で、私たちは、(I)1つ1つのRNA結合蛋白質に焦点をあて脳組織、あるいは細胞からマルチオミクス解析により分子精査することで、その機能の解明を目指す研究を行ってきた。(II)同時に、膨大なRNA結合蛋白質群のうち、どのRNA結合因子に着目するべきかという問いに対し、RNA結合蛋白質群の分類学的思考を凝らした包括的解析を試みてきた。
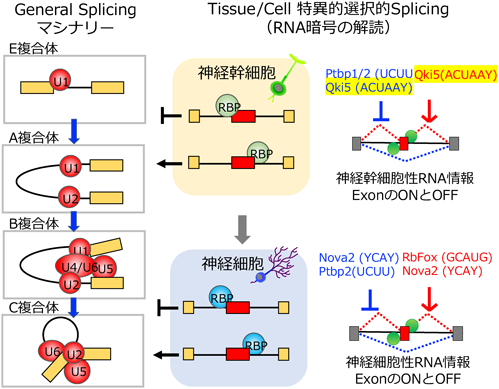
詳細は、他書へ譲りますが、はじめに私が記した2004年にコールドスプリングハーバー研究所で見たオブジェの夢に答える研究手法がHITS-CLIP法であります5–7)。大袈裟に言うと本手法を、脳細胞機能の解析に世界で初めて導入した仕事が、私の留学時代の研究成果であります。大学院で学んだ神経発生学、ニューロサイエンスの知識を活かし、蛋白質–RNA相互作用マッピング技術HITS-CLIPを用いて、神経特異的RNA結合蛋白質Nova2がDab-1シグナリングを介した大脳皮質の神経細胞移動に働くことを明らかにした8)。実際、Nova2欠損マウスは、大脳皮質および小脳プルキンエ細胞の神経細胞移動に対し、リーリンシグナル異常と類似した表現型を示す8, 9)。次に、帰国後、自身の研究チームとして、院生時代当初の願望である神経発生に関わる重要分子の発見というキーワードのもとで、私たちは、マウスの胎児期において、神経幹細胞に限局して発現するRNA結合蛋白質Qki5の機能解析の着手した。Qki5は、神経幹細胞の神経分化のごく初期のタイミングを担うTbr2陽性細胞で、発現がシャットアウトする分子であることを見出した。神経発生学での知恵では、まさに幹細胞性に関わる分子として期待され、私たちにとって興味深いものであった。実際、遺伝子操作や遺伝子改変動物を用いたトランスクリプトーム解析とHITS-CLIP技術を組み合わせた研究戦略により、850程度の標的RNA群を同定し、特にβ-cateninシグナルを介した神経幹細胞性を制御する因子として論文を発表した10)。特に、HITS-CLIP技術は、さらに進化を遂げ、1塩基解像度でQki5のRNA結合部位をトランスクリプトームワイドで明らかにすることができた10, 11)(図1右)。ここまでの神経発生に関わるRNA結合蛋白質群の解析を合わせ、神経幹細胞–神経細胞分化遷移に働くRNA結合部位依存的な選択的スプライシング制御のRNA暗号の解読の成功として纏められる10, 12–14)。実際、最新のこの領域の総説を読みながら本分野への貢献を感じとることができる15)(図2)。
RNA結合蛋白質、RNA関連の疾患研究が盛んになる中で、癌や筋萎縮性側索硬化症(ALS)、22q11.2欠失症候群などの疾患モデルを確立することが重要であり、幸運にもこれらモデルを用いた共同研究に、特にRNA解析を中心に携わる機会を得た16–18)。ところが解析の実際は、RNA結合蛋白質の下流RNAの制御が“ファインチューニング”などと言われる通り、所謂、遺伝子発現の柔軟性を制御するもので、単一分子経路に行き着くのが困難で多くの試行錯誤を要した。例えば、22q11.2欠失症候群では、数ある遺伝子領域のマイクロ欠失によるハプロ不全症の疾患であり、その候補原因遺伝子がmiRNAの生合成を司るマイクロプロセッサー複合体のコア分子、二本鎖RNA結合蛋白質DGCR8と考えられている。miRNA合成のコア分子であることから全てのmiRNA合成に必須である一方で、ハプロ不全による活性が半減した状態では、miRNA種ごとにその感受性が異なるというミステリーがあった19)。そこで私たちは、特に神経系で発現し、かつDGCR8感受性の高いmiR-9-2に着目してdualカラーのmiRNAプロセッサーレポーター解析系を確立し、新規のDGCR8作動エレメントDREの同定とその活性にRNA修飾が寄与していることを見出す事ができた20)。本レポーターモデルは、RNA結合蛋白質と標的RNAの機能解析について、miRNAのプロセッシングだけでなく、スプライシングや翻訳制御さらに化合物スクリーニングも可能なツールとなった。
RNA結合蛋白質は、細胞の生存そのものに必須な基本マシナリーに働く分子から、細胞種、組織特異的に働くもの、一見必要なさそうな分子(厳密に言うとそんな分子はない!)まで幅広く存在する。その中で、疾患や発生に関わる重要なRNA結合蛋白質を抽出し、上記で述べた1つのRNA結合蛋白質の解析へとつなげる手段として、特定の疾患、発生段階における細胞種にエンリッチして発現するRNA結合蛋白質の同定、あるいは、病態のトランスクリプトーム情報に対して強いインパクト、影響力を示すRNA結合蛋白質の階層クラス分けを試みた。特に、FUS-ALSの細胞モデルを確立したことで17)、運動ニューロンへの分化段階、FUS変異をホモに持つもの、ヘテロに持つものなど計60種類の様々なトランスクリプトーム情報に内在するRNA制御因子群のヒエラルキーをベイジアンネットワーク解析によって明らかにする試みを行った。当時は、まだ斬新なアイデアだったと思うのだが、同じくAIを用いた解析、例えば2018年に報告された同様の試みとしては、IBM-ワトソン君による文献駆動型(Literature-driven)解析で、ALSに関わるRNA結合蛋白質としてHNRNPUがピックアップされている21)。奇しくも、我々もHNRNPUのOptimized HITS-CLIP解析で炎症性サイトカインIL-6産生に関わる分子としてHNRNPUの論文発表を行なっている11)。一方、私たちの解析では、完全データ駆動型解析(Data-driven)で、iBRN(iPS由来神経細胞を使ったベイジアンネットワーク)解析と名付けて、紆余曲折、初投稿から3年以上の歳月をかけて無事に昨年、世に公表することができた22)。病態のRNA情報に対して、RNA制御因子群の上下関係を図3のように推定するもので、上位因子ほど、子供因子を多く持つ親分子となり、病態に対してハブ因子(hub RBP)と名付け、想定するものである(図3)。実際、既報のFUS蛋白質の液相分離–固相分離を制御するPRKDCがHub RBPとして含まれるもので、結果はとても納得のいく素晴らしいものであった。また、RNA発現情報のみを指標とするものであるため、機能未知な非コードRNAなども解析の対象となるのが魅力的である。実際、遺伝子解析で候補ハブ因子を操作することで、DNA損傷刺激が誘発されるなど、神経変性疾患へと誘導される経路が活性化された22)。さらに、候補ハブ因子となるRNA制御因子の解析を継続していると大変興味深い現象に気づかされる。それは、ハブ因子が、DNA損傷、概日リズム、炎症反応、非コードRNA制御といった疾患へと至るような複数の分子経路の階層を跨ぐ超階層性ネットワークを有するということである(図3右)。
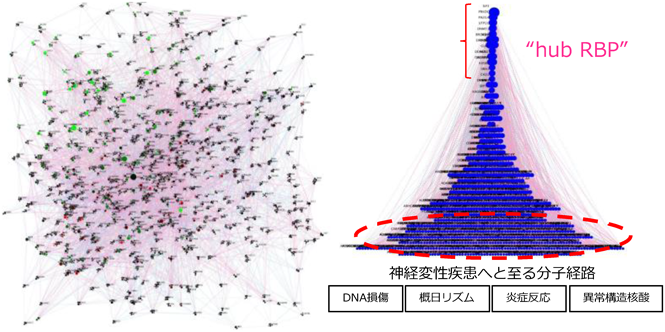
ビジネスや社会学、組織論で耳にする超階層性ネットワークという言葉があるが、RNA結合蛋白質による制御モデルはそれにとてもフィットしうると考えられる。学生時代に頭の中で整理していた特徴的な発現パターンを有する神経発生学の本質を担う転写因子群は、どちらかというと明確かつ強力な命令系統を持つマスター制御因子の1つだと捉えている。一方で、トランスクリプトーム情報に対してインパクトを示すハブRNA結合蛋白質は、発生分化、細胞老化、炎症反応、細胞機能に対して超階層的な指示系統を持つ一方で、いずれも緩やかかつ多岐にわたる指示系統分子と言える。RNA結合蛋白質群は、まさに、近年、語られるロバストネス、生老病死を回避するための脳機能の柔軟性を司る分子群として今後も益々興味深い研究対象である。
これまで、一貫してRNA結合蛋白質が非常に面白いと思って研究してきたが、今回、思い返してみると自身の研究の進んできた軌道修正が興味深い。無論、技術的進歩によるものであるが、技術そのものの更新も当然、必要であり、まして観察結果の考察や仮説の更新は常に必要であるのは言うまでもない。研究の醍醐味は、「わかった、理解した」にあり、とても嬉しい事である。それでも、これからも「本当にわかったの?」を大切に常に意識し、新たな疑問や仮説を更新しながら研究してみたい。それには、今後もRNA結合蛋白質群の分類、そして1つのRNA結合蛋白質解析への注力、さらに柔軟な脳組織が機能しうる超階層性ネットワークの理解という無限ループの考察、更新を続け、できる限り本研究を続ける事が私の現地点での目標である。
本研究を遂行する上でご指導頂いた恩師、共同研究していただいた多くの先生方にこの場を借りて感謝致します。また、本稿執筆にあたり、朝から晩までRNA結合蛋白質について議論してくれた仲間たち、産学連携研究の共同研究者、野上真宏博士、湯上真人博士、松尾毅博士と矢野(早川)佳芳博士、古川貴子さん、小山哲秀博士には特別の感謝を伝えたい。最後に本執筆の機会をいただいた等誠司先生、竹林浩秀先生、そして芝田晋介先生に心より深謝致します。
1) Castello A, Fischer B, Eichelbaum K, Horos R, Beckmann BM, Strein C, Davey NE, Humphreys DT, Preiss T, Steinmetz LM, Krijgsveld J, Hentze MW. Insights into RNA biology from an atlas of mammalian mRNA-binding proteins. Cell, 149(6), 1393–1406 (2012). doi: 10.1016/j.cell.2012.04.031.
2) Gerstberger S, Hafner M, Tuschl T. A census of human RNA-binding proteins. Nat Rev Genet, 15(12), 829–845 (2014). doi: 10.1038/nrg3813.
3) Yano M, Okano HJ, Okano H. Involvement of Hu and heterogeneous nuclear ribonucleoprotein K in neuronal differentiation through p. 21 mRNA post-transcriptional regulation. J Biol Chem, 280(13), 12690–12699 (2005). doi: 10.1074/jbc.M411119200.
4) Akamatsu W, Fujihara H, Mitsuhashi T, Yano M, Shibata S, Hayakawa Y, Okano HJ, Sakakibara S, Takano H, Takano T, Takahashi T, Noda T, Okano H. The RNA-binding protein HuD regulates neuronal cell identity and maturation. Proc Natl Acad Sci USA, 102(12), 4625–4630 (2005). doi: 10.1073/pnas.0407523102.
5) Licatalosi DD, Mele A, Fak JJ, Ule J, Kayikci M, Chi SW, Clark TA, Schweitzer AC, Blume J, Wang X, Darnell JC, Darnell RB. HITS-CLIP yields genome-wide insights into brain alternative RNA processing. Nature, 456(7221), 464–469 (2008). doi: 10.1038/nature07488.
6) Licatalosi DD, Darnell RB. RNA processing and its regulation: global insights into biological networks. Nat Rev Genet, 11(1), 75–87 (2010). doi: 10.1038/nrg2673.
7) 矢野真人,矢野佳芳,Robert Darnell.クローズアップ実験法Series210 HITS-CLIP法:タンパク質–RNA相互作用のゲノムワイドマッピング.実験医学,105–114 (2011).
8) Yano M, Hayakawa-Yano Y, Mele A, Darnell RB. Nova2 regulates neuronal migration through an RNA switch in disabled-1 signaling. Neuron, 66(6), 848–858 (2010). doi: 10.1016/j.neuron.2010.05.007.
9) Hack I, Hellwig S, Junghans D, Brunne B, Bock HH, Zhao S, Frotscher M. Divergent roles of ApoER2 and Vldlr in the migration of cortical neurons. Development, 134(21), 3883–3891 (2007). doi: 10.1242/dev.005447.
10) Hayakawa-Yano Y, Suyama S, Nogami M, Yugami M, Koya I, Furukawa T, Zhou L, Abe M, Sakimura K, Takebayashi H, Nakanishi A, Okano H, Yano M. An RNA-binding protein, Qki5, regulates embryonic neural stem cells through pre-mRNA processing in cell adhesion signaling. Genes Dev, 31(18), 1910–1925 (2017). doi: 10.1101/gad.300822.117.
11) Yugami M, Okano H, Nakanishi A, Yano M. Analysis of the nucleocytoplasmic shuttling RNA-binding protein HNRNPU using optimized HITS-CLIP method. PLoS One, 15(4), e0231450 (2020). doi: 10.1371/journal.pone.0231450.
12) Licatalosi DD, Yano M, Fak JJ, Mele A, Grabinski SE, Zhang C, Darnell RB. Ptbp2 represses adult-specific splicing to regulate the generation of neuronal precursors in the embryonic brain. Genes Dev, 26(14), 1626–1642 (2012). doi: 10.1101/gad.191338.112.
13) Yano M, Ohtsuka T, Okano H. RNA-binding protein research with transcriptome-wide technologies in neural development. Cell Tissue Res, 359(1), 135–144 (2015). doi: 10.1007/s00441-014-1923-8.
14) Hayakawa-Yano Y, Yano M. An RNA switch of a large exon of Ninein is regulated by the neural stem cell specific-RNA binding protein, Qki5. Int J Mol Sci, 20(5), 1010 (2019). doi.org/10.3390/ijms20051010.
15) Fisher E, Feng J. RNA splicing regulators play critical roles in neurogenesis. Wiley Interdiscip Rev RNA, e1728 (2022).
16) Toyoshima M, Akamatsu W, Okada Y, Ohnishi T, Balan S, Hisano Y, Iwayama Y, Toyota T, Matsumoto T, Itasaka N, Sugiyama S, Tanaka M, Yano M, Dean B, Okano H, Yoshikawa T. Analysis of induced pluripotent stem cells carrying 22q11.2 deletion. Transl Psychiatry, 6(11), e934 (2016). doi: 10.1038/tp.2016.206.
17) Ichiyanagi N, Fujimori K, Yano M, Ishihara-Fujisaki C, Sone T, Akiyama T, Okada Y, Akamatsu W, Matsumoto T, Ishikawa M, Nishimoto Y, Ishihara Y, Sakuma T, Yamamoto T, Tsuiji H, Suzuki N, Warita H, Aoki M, Okano H. Establishment of in vitro FUS-associated familial amyotrophic lateral sclerosis model using human induced pluripotent stem cells. Stem Cell Reports, 6(4), 496–510 (2016). doi: 10.1016/j.stemcr.2016.02.011.
18) Fox RG, Lytle NK, Jaquish DV, Park FD, Ito T, Bajaj J, Koechlein CS, Zimdahl B, Yano M, Kopp J, Kritzik M, Sicklick J, Sander M, Grandgenett PM, Hollingsworth MA, Shibata S, Pizzo D, Valasek M, Sasik R, Scadeng M, Okano H, Kim Y, MacLeod AR, Lowy AM, Reya T. Image-based detection and targeting of therapy resistance in pancreatic adenocarcinoma. Nature, 534(7607), 407–411 (2016). doi: 10.1038/nature17988.
19) Stark KL, Xu B, Bagchi A, Lai WS, Liu H, Hsu R, Wan X, Pavlidis P, Mills AA, Karayiorgou M, Gogos JA. Altered brain microRNA biogenesis contributes to phenotypic deficits in a 22q11-deletion mouse model. Nat Genet, 40(6), 751–760 (2008). doi: 10.1038/ng.138.
20) Nogami M, Miyamoto K, Hayakawa-Yano Y, Nakanishi A, Yano M, Okano H. DGCR8-dependent efficient pri-miRNA processing of human pri-miR-9-2. J Biol Chem, 296, 100409 (2021). doi: 10.1016/j.jbc.2021.100409.
21) Bakkar N, Kovalik T, Lorenzini I, Spangler S, Lacoste A, Sponaugle K, Ferrante P, Argentinis E, Sattler R, Bowser R. Artificial intelligence in neurodegenerative disease research: use of IBM Watson to identify additional RNA-binding proteins altered in amyotrophic lateral sclerosis. Acta Neuropathol, 35(2), 227–247 (2018). doi: 10.1007/s00401-017-1785-8.
22) Nogami M, Ishikawa M, Doi A, Sano O, Sone T, Akiyama T, Aoki M, Nakanishi A, Ogi K, Yano M, Okano H. Identification of hub molecules of FUS-ALS by Bayesian gene regulatory network analysis of iPSC model: iBRN. Neurobiol Dis, 155, 105364 (2021). doi: 10.1016/j.nbd.2021.105364.

This page was created on 2022-06-06T13:02:43.126+09:00
This page was last modified on 2022-07-21T15:27:11.000+09:00
このサイトは(株)国際文献社によって運用されています。