生体高分子のリソソームへの新たな輸送・分解経路
1 大阪大学大学院 連合小児発達学研究科 分子生物遺伝学研究領域
2 国立精神・神経医療研究センター 神経研究所 疾病研究第四部
発行日:2022年6月30日Published: June 30, 2022
© 2022 日本神経化学会© 2022 The Japanese Society for Neurochemistry
生命は様々な生体物質の合成と分解のバランスの上に成り立っており、恒常性の維持において細胞内構成因子の適切な分解は必須である。核酸やタンパク質、脂質などの生体高分子は細胞内で絶えず合成されており、これらの分解が滞ることが多様な疾患の原因となることが知られている。リソソームは細胞内最大の物質分解の場であり、生体高分子のリソソーム性分解の破綻は多くの場合で神経症状を示す様々な疾患の原因となる1)。また神経変性疾患や一部のミオパチーにおいては細胞内に凝集性タンパク質等、様々な物質の蓄積が見られ、細胞内分解系の破綻が病態の形成に深く関わっていることが示唆されてきた。
リソソームに基質となる細胞内物質を輸送し分解する機構を広義にオートファジーと総称する。オートファジーにはこれまで大きく分けてマクロオートファジー、ミクロオートファジー、シャペロン介在性オートファジーの3つの経路が知られていた2)。これまでのオートファジー研究は、マクロオートファジー(狭義の「オートファジー」)研究を中心に本邦が世界をリードし精力的に推進されてきたが、マクロオートファジー以外の経路については、相対的に研究が立ち遅れていた。加えて既知の細胞内分解経路のみでは細胞内物質分解の多くの割合を説明できないとの指摘もあり3)、未知のシステムの存在も示唆されていた。
このような背景の下、筆者は細胞内物質分解システムの全容解明を目標に、リソソームへの新たな分解基質輸送経路の探索を行ってきた。筆者らはこれまでに「リソソームがATP依存的に核酸やタンパク質を直接取り込み、分解する」という新たな細胞内物質輸送・分解経路を見出し、その分子機構や生理的・病態生理的意義を探求してきた。本稿では筆者らがdirect uptake via/through membrane protein(DUMP)と名づけたこの細胞内分解システム(図1)について、これまでに明らかとなってきたことや、最新の知見について概説したい。
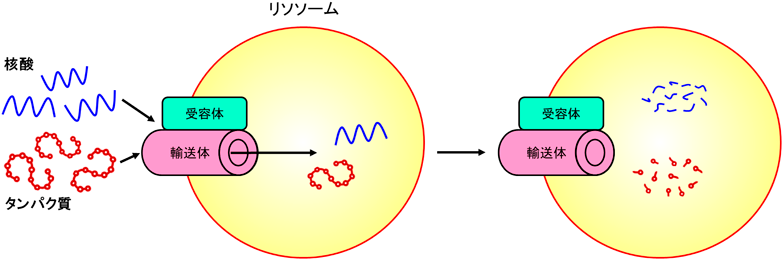
リソソームは1955年にChristian de Duveらによって報告された4)、内部に多様な加水分解酵素を含むオルガネラであり、ほぼすべての生体高分子を分解することができる唯一の細胞内器官である5)。この直径数百ナノメートルのコンパートメントの内腔に、現在知られているだけで50種類以上もの加水分解酵素が存在する6)。リソソームに細胞内の物質を輸送し分解する機構を広義にオートファジーと総称するが、「オートファジー」という用語もまた、細胞外の物質をリソソームへと運ぶ「ヘテロファジー」(エンドサイトーシスやファゴサイトーシス)と対になる語として、de Duveによって提唱されたものである。オートファジーにはこれまで、大きく分けてマクロオートファジー、ミクロオートファジー、シャペロン介在性オートファジー(CMA)の3つの経路が知られていた。これらの中で圧倒的に研究が進んでいるのはマクロオートファジーであり、通常、単にオートファジーと述べた場合、狭義にマクロオートファジーのことを指すのはひとつにはこのためである。マクロオートファジーは物質輸送過程にオートファゴソームと呼ばれる、リソソームとは別の小胞による輸送を介すシステムであるが、他の2経路はリソソーム自体が分解基質を取り込む経路であり、これらを含むリソソームそのものによる物質輸送機構に関してはいまだその分子機構に不明な点や、また未知の経路の残されている可能性もあり、未開拓・発展途上な部分が多いのが実情である。
内腔に存在するさまざまな加水分解酵素に加えてリソソームにはもうひとつ、豊富かつ多様な膜タンパク質を有するという特徴がある。研究にもよるが、およそ25種類前後のリソソーム膜タンパク質がこれまでに知られており6)、いまだ多くの機能未知のものが残されている。筆者らは当初、このような機能未知のリソソーム膜タンパク質の中に、何らかの分解基質の取り込みに関わる因子が存在するのではないかと考え研究を開始した。
この研究において筆者らはまずLysosome-associated membrane protein type 2(LAMP2)と呼ばれるリソソーム膜タンパク質に注目した。LAMP2は1回膜貫通型のタンパク質で、リソソームで最も豊富な膜タンパク質のひとつである。また、LAMP2の内腔ドメインは高度に糖鎖修飾を受けており、これらの特徴から古くはリソソーム内腔の加水分解酵素から膜を保護し、リソソーム自体の崩壊を防ぐ、構造タンパク質だと考えられてきた7)。しかしながらLAMP2にはその構造の大部分を占める内腔ドメインが共通でありながら、ごく短い膜貫通~細胞質側配列が異なるA、B、Cというスプライスバリアントが知られていた7)。このようなバリアントの存在意義を前述の構造タンパク質としての側面だけで説明するのは難しい。加えてLAMP2の中でLAMP2Aのみがその固有の細胞質側配列を介して、既知のリソソームへのタンパク質直接取り込み経路、シャペロン介在性オートファジーの基質受容体として働くことが知られていた8)。我々はLAMP2BやCにも、個別の細胞質側配列に結合する分子が存在し、それこそがこれらLAMP2バリアントの存在意義なのではないかと考えた。そこでまずそれぞれの細胞質側配列に結合する生体高分子を探索したところ、LAMP2Cの細胞質側配列にRNAが特異的に直接結合することを見出した9)。それまでリソソーム内部に局在するRNAやDNAの分解酵素は古くから知られていた一方で、核酸を直接の標的とするオートファジー機構は知られていなかった10)。以上より筆者らはシャペロン介在性オートファジーにおけるLAMP2Aのように、LAMP2Cを受容体としてRNAを直接リソソームへと取り込むような経路が存在するのではないかと推測し、まず、そもそもリソソームがRNAを直接内腔へと運び込むことができるのか、検討を行った。
物質のオルガネラへの取り込みが「直接」であることを実験的に示すには、他の介在要素の不在を担保する必要がある。筆者はこの点を克服するため、リソソームを細胞や組織から単離し、ATPの存在下あるいは非存在下において基質の精製RNAと37°Cでインキュベートすることでリソソームによる直接的なRNA取り込みや分解を観察する、in vitro再構成実験系を構築した9)。その結果、リソソームとRNAをそのままインキュベートしても明らかな変化は見られない一方で、ATPを添加するとリソソームがRNAを直接取り込み、分解することが、生化学的解析や免疫電子顕微鏡法を用いた観察により明らかとなった9)。さらにこの活性はLAMP2Cの過剰発現およびLAMP2のノックアウト(KO)によってそれぞれ増強・減弱したことから、少なくとも一部においてLAMP2Cに依存していることもわかった9)。細胞レベルでの解析でも、LAMP2Cの過剰発現で細胞全体における総RNA分解量の上昇が見られ、またLAMP2 KOマウスでは老齢期の脳におけるRNAの有意な蓄積が見られた。脳、その中でも神経細胞はLAMP2Cの発現が特に高い組織・細胞である。これらの傍証から、生きた細胞や動物個体においても同様の現象が起きていることが示唆される9)。以上に加えて筆者らはDNAがLAMP2Cの細胞質側配列に結合し、少なくともin vitroにおいてRNAと同様にリソソームによりATP依存的に直接取り込まれ、分解を受けることを見出し、報告している11)。
LAMP2の細胞質側配列の長さはいずれのバリアントも11-12アミノ酸であり、当初このごく短い配列にどのようにして核酸が結合するのかは不明であった。筆者らはLAMP2Cの細胞質側配列の中にarginine-rich motif(ARM)と呼ばれる古くから知られる核酸結合モチーフが存在し、この配列を介してRNAやDNAが結合していることを見出した12)。線虫(C. elegans)やショウジョウバエ(D. melanogaster)では脊椎動物のようにLAMPにバリアントが存在しないが、興味深いことにこれらの細胞質側配列はいずれもヒトLAMP2の中でバリアントCのものに最も高い相同性を示し、かつ核酸結合能を持っていた9, 12)。このことはARMを介した核酸結合が進化的にLAMPで保存されてきた古い機能である可能性を示唆する。このような配列的特徴は脊椎動物では部分的にLAMP2Bや他のLAMPファミリータンパク質にも共有されており、これらの一部においてもLAMP2Cの細胞質側配列よりは弱い核酸結合能が認められた12)。ARMではアルギニンとリジン残基が核酸結合において重要な役割を果たすが、前述のシャペロン介在性オートファジーの受容体LAMP2Aの細胞質側配列からはアルギニン残基が排除されており、かわりに同じ塩基性アミノ酸であるヒスチジンの連続配列とリジン残基が基質タンパク質との結合に関わる点は興味深い8)。
続いて筆者らはLAMP2Cによって認識される核酸の配列についても検討を行ったところ、人工的に合成した単一塩基の連続配列の中ではRNA、DNAともにグアニンの連続配列のみが結合性を示し、リソソームへの取り込みについてもこれらのみが確認された13)。これらはあくまで単一の塩基から成る人工的な配列についての検討であるが、核酸のLAMP2C細胞質側配列に対する結合の有無とリソソームへの取り込みが相関することは、本経路における基質と受容体の結合が重要なステップであることを示唆する。
ここまでの研究でリソソームへの新たな核酸取り込み・分解経路とその核酸受容体LAMP2Cの存在が示されたものの、本研究には開始当初からの疑問点が残されていた。核酸のような親水性の分子が限界膜を通過するためには通常、何らかの輸送体を必要とするが、輸送体として働くタンパク質は一般に複数回膜貫通型の構造を持つ14)。前述の通りLAMP2Cは一回膜貫通型のタンパク質であり、この点に乖離があった。なお、シャペロン介在性オートファジーでは例外的にLAMP2Aが基質タンパク質の取り込み過程までも担うというモデルが提唱されているが、その輸送実体に対しては「translocation complex(移行複合体)」という慎重な呼称が用いられている15)。筆者らはLAMP2Cとは別に、核酸のリソソームへの取り込みをより直接的に担う因子が存在するのではないかと類推していた。これに対し筆者らは9回膜貫通型リソソーム膜タンパク質SID1 transmembrane family member 2(SIDT2)がリソソームへの核酸の直接取り込みを担うことを見出した16, 17)。SIDT2は線虫において細胞膜の核酸トランスポーターとして報告されているsystemic RNA interference defective protein 1(SID1)18, 19)の哺乳類オルソログのひとつであり、リソソーム膜局在を示す。筆者らはこのSIDT2こそが本経路における核酸の輸送実体なのではないかと推測し、まずここまでの研究で確立していたin vitro再構成実験系を用いてリソソームへのRNA直接取り込みに対するSIDT2の関与を検討した。その結果、やはりSIDT2過剰発現細胞由来のリソソームでは取り込み・分解活性の増強が、SIDT2ノックダウン(KD)細胞由来のリソソームでは減弱がそれぞれ見られた16)。また、DNAの取り込み・分解についても同様のSIDT2の効果が見られている17)。細胞レベルでの検討においても、SIDT2 KDマウス胎児線維芽細胞で細胞内総RNA分解量に対し最大で約50%の低下が見られた16)。またSIDT2の過剰発現によっても細胞内総RNA分解量の顕著な増加が見られている20)。これらに加えて、先述の線虫SID1に関する先行研究ではRNA輸送活性に必須なセリン残基が特定されており18)、この残基はSIDT2においても保存されていた。そこで当該のセリン残基を置換したSIDT2変異体の活性を調べたところ、このSIDT2変異体はRNA、DNAいずれのリソソームへの取り込みに対しても機能しないことが明らかとなった16, 17)。これらより、核酸のリソソームへの直接取り込みを担う重要な輸送体の少なくともひとつがSIDT2であると考えられる。このSIDT2のリソソーム膜局在については、アミノ酸配列中に3箇所のリソソーム局在シグナルが存在し、これらの配列を置換したSIDT2はリソソーム局在を失うとともに核酸の取り込み・分解促進活性も失うことから、そのいずれもがSIDT2のリソソーム膜への適切な局在化に重要であることも明らかとなっている20)。また、興味深いことに筆者らはSIDT2の細胞質側に露出した箇所の配列にはLAMP2Cと同様、先述の核酸結合モチーフARMが存在し、この配列がSIDT2の核酸との結合とリソソームへの取り込みいずれにおいても必要であることも見出した21)。これはすなわちSIDT2がLAMP2Cと同様の核酸結合様式によって機能することことを意味する。LAMP2CはSIDT2の機能において必須ではないものの両者には相互作用が見られ、かつSIDT2とLAMP2Cには核酸の分解に対する相乗的な関係が見られたことなどから、SIDT2の核酸取り込み活性に対しLAMP2Cは受容体としての補助的役割を担うと考えられる16, 21)。
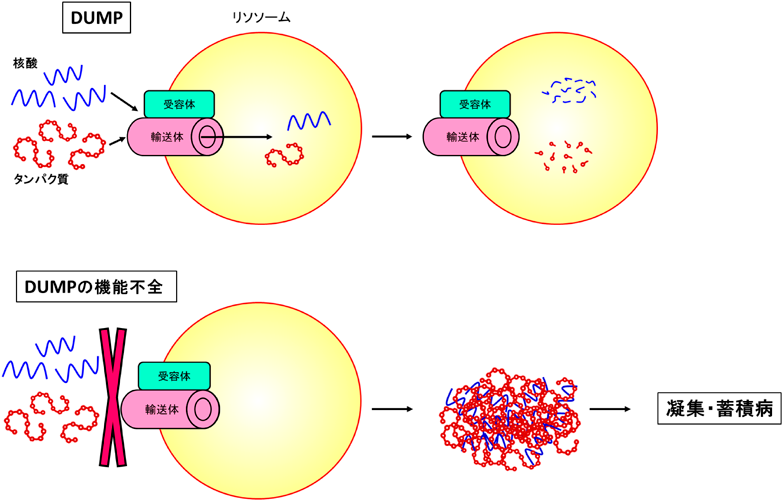
ここまでで紹介した研究の過程において筆者らは最近、SIDT2がどうやら核酸だけでなく細胞内のタンパク質の分解にも深く関わるようだということに気づいた。培養細胞ではSIDT2の増減がリソソーム性の細胞内タンパク質分解量を変化させ、単離リソソームを用いた実験からSIDT2がタンパク質のリソソームへの直接取り込み・分解を仲介することが明らかとなった。筆者らは改めてこのようなリソソームによる核酸やタンパク質など多様な生体高分子の直接取り込み経路を“direct-uptake-via/through-membrane-protein”(DUMP)と新たに名づけ、現在論文のプレプリントを公開中である22)。さらにこの研究において筆者らはDUMPの機能不全を原因とすると考えられる家族性神経筋疾患を1例、見出すに至った(図2)22)。この疾患患者はSIDT2にDUMPの活性低下を惹き起こす変異を持っており、患者筋生検においては凝集性タンパク質等、様々な物質の細胞質への蓄積が見られた。このような蓄積・凝集物や筋病理所見を含め、SIDT2 KOマウスにおいて患者と非常によく似た表現型が得られていることも、DUMPの活性低下がこのような疾患の原因となっていることを支持する22)。これに続き筆者らはパーキンソン病やレビー小体型認知症患者の死後脳における、SIDT2の病態依存的な発現上昇や局在変化を見出している23)。これらの疾患の患者死後脳(前帯状皮質)においてはSIDT2の増加がいずれも見られ、さらにコントロール例も含めたすべてのサンプルにおいて脳内α-synucleinタンパク質の量とSIDT2量に強い相関が見られた23)。α-synucleinタンパク質はこれらレビー小体病において凝集・蓄積する主要因子であり、またDUMPの分解基質となること22)、老齢SIDT2欠損マウス脳ではα-synucleinの蓄積が見られること23)などから、これらの疾患の病態形成過程におけるα-synucleinの増加に対する恒常性維持機構としてSIDT2が発現上昇している可能性が考えられる。一方で患者死後脳の免疫染色ではSIDT2のリン酸化α-synuclein凝集物への集積が見られたことから23)、SIDT2やそれが局在するリソソームがレビー小体へとトラップされることによるDUMP活性の低下が病態進行の一因となっている可能性も考えられ24)、さらなる解析が必要である。
細胞核の発見が1831年、ミトコンドリアの発見が1897年であることを考慮すると、リソソームは比較的「若い」オルガネラであると言えるかもしれない。Rudolph Schoenheimerが細胞内分子の絶え間ない合成と分解の両輪から成る生体物質の入れ替わりの概念を提唱したのは1930–40年代のことであるが、その後のセントラルドグマの提唱と分子生物学の勃興の中で生命科学の中心をまず担ったのは細胞内合成系に関する研究であった。オートファジーの中で圧倒的に研究の進んでいるマクロオートファジーに関する研究が花開いたのも、1990年代からのことである25)。本稿で紹介させていただいた内容も含め、リソソームにはいまだ知られていなかった機能が残されており、また今後も未知の経路や機能が発見される可能性も大いにあると考えられる。冒頭でも述べた通り神経や筋はオートファジー・リソソーム系等、細胞内分解系の破綻の影響が出やすい組織であり、その解明は神経化学や医学の発展にとっても重要な意義を持ちうるものである。
本研究を行うにあたり、多大なるご指導とご協力を賜りました国立精神・神経医療研究センター神経研究所の和田圭司先生、株田智弘先生、研究室の皆様、多くの共同研究者の先生方にこの場をお借りして心より感謝申し上げます。また、この度このように光栄な場での執筆の機会を与えてくださいました日本神経化学会出版・広報委員会の等誠司委員長、委員の先生方ならびに編集部の皆様に深く御礼申し上げます。
1) Klein AD, Futerman AH. Lysosomal storage disorders: Old diseases, present and future challenges. Pediatr Endocrinol Rev, 11(Suppl 1), 59–63 (2013).
2) Mizushima N, Komatsu M. Autophagy: renovation of cells and tissues. Cell, 147(4), 728–741 (2011).
3) Tong M, Smeekens JM, Xiao H, Wu R. Systematic quantification of the dynamics of newly synthesized proteins unveiling their degradation pathways in human cells. Chem Sci (Camb), 11(13), 3557–3568 (2020).
4) de Duve C, Pressman BC, Gianetto R, Wattiaux R, Appelmans F. Tissue fractionation studies. 6. Intracellular distribution patterns of enzymes in rat-liver tissue. Biochem J, 60(4), 604–617 (1955).
5) Mizushima N. A brief history of autophagy from cell biology to physiology and disease. Nat Cell Biol, 20(5), 521–527 (2018).
6) Saftig P, Klumperman J. Lysosome biogenesis and lysosomal membrane proteins: Trafficking meets function. Nat Rev Mol Cell Biol, 10(9), 623–635 (2009).
7) Eskelinen EL, Cuervo AM, Taylor MR, Nishino I, Blum JS, Dice JF, Sandoval IV, Lippincott-Schwartz J, August JT, Saftig P. Unifying nomenclature for the isoforms of the lysosomal membrane protein LAMP-2. Traffic, 6(11), 1058–1061 (2005).
8) Cuervo AM, Dice JF. Unique properties of lamp2a compared to other lamp2 isoforms. J Cell Sci, 113(Pt 24), 4441–4450 (2000).
9) Fujiwara Y, Furuta A, Kikuchi H, Aizawa S, Hatanaka Y, Konya C, Uchida K, Yoshimura A, Tamai Y, Wada K, Kabuta T. Discovery of a novel type of autophagy targeting RNA. Autophagy, 9(3), 403–409 (2013).
10) Fujiwara Y, Wada K, Kabuta T. Lysosomal degradation of intracellular nucleic acids—multiple autophagic pathways. J Biochem, 161(2), 145–154 (2017).
11) Fujiwara Y, Kikuchi H, Aizawa S, Furuta A, Hatanaka Y, Konya C, Uchida K, Wada K, Kabuta T. Direct uptake and degradation of DNA by lysosomes. Autophagy, 9(8), 1167–1171 (2013).
12) Fujiwara Y, Hase K, Wada K, Kabuta T. An RNautophagy/DNautophagy receptor, LAMP2C, possesses an arginine-rich motif that mediates RNA/DNA-binding. Biochem Biophys Res Commun, 460(2), 281–286 (2015).
13) Hase K, Fujiwara Y, Kikuchi H, Aizawa S, Hakuno F, Takahashi S, Wada K, Kabuta T. RNautophagy/DNautophagy possesses selectivity for RNA/DNA substrates. Nucleic Acids Res, 43(13), 6439–6449 (2015).
14) Alberts B, Johnson A, Lewis J, Raff M, Roberts K, Walter P.細胞の分子生物学,651–694, Newton Press, (2010).
15) Kaushik S, Cuervo AM. The coming of age of chaperone-mediated autophagy. Nat Rev Mol Cell Biol, 19(6), 365–381 (2018).
16) Aizawa S, Fujiwara Y, Contu VR, Hase K, Takahashi M, Kikuchi H, Kabuta C, Wada K, Kabuta T. Lysosomal putative RNA transporter SIDT2 mediates direct uptake of RNA by lysosomes. Autophagy, 12(3), 565–578 (2016).
17) Aizawa S, Contu VR, Fujiwara Y, Hase K, Kikuchi H, Kabuta C, Wada K, Kabuta T. Lysosomal membrane protein SIDT2 mediates the direct uptake of DNA by lysosomes. Autophagy, 13(1), 218–222 (2017).
18) Shih JD, Hunter CP. SID-1 is a dsRNA-selective dsRNA-gated channel. RNA, 17(6), 1057–1065 (2011).
19) Xu J, Yoshimura K, Mon H, Li Z, Zhu L, Iiyama K, Kusakabe T, Lee JM. Establishment of Caenorhabditis elegans SID-1-dependent DNA delivery system in cultured silkworm cells. Mol Biotechnol, 56(3), 193–198 (2014).
20) Contu VR, Hase K, Kozuka-Hata H, Oyama M, Fujiwara Y, Kabuta C, Takahashi M, Hakuno F, Takahashi SI, Wada K, Kabuta T. Lysosomal targeting of SIDT2 via multiple YxxΦ motifs is required for SIDT2 function in the process of RNautophagy. J Cell Sci, 130(17), 2843–2853 (2017).
21) Hase K, Contu VR, Kabuta C, Sakai R, Takahashi M, Kataoka N, Hakuno F, Takahashi SI, Fujiwara Y, Wada K, Kabuta T. Cytosolic domain of SIDT2 carries an arginine-rich motif that binds to RNA/DNA and is important for the direct transport of nucleic acids into lysosomes. Autophagy, 16(11), 1974–1988 (2020).
22) Fujiwara Y, Contu VR, Kabuta C, Ogawa M, Fujita H, Kikuchi H, Sakai R, Hase K, Suzuki M, Koyama-Honda I, Inoue M, Oya Y, Inoue YU, Inoue T, Takahashi R, Nishino I, Wada K, Noguchi S, Kabuta T. Discovery of a protein uptake pathway in lysosomes. bioRxiv, 2020.2008.2011.245688 (2021).
23) Fujiwara Y, Kabuta C, Sano T, Murayama S, Saito Y, Kabuta T. Pathology-associated change in levels and localization of SIDT2 in postmortem brains of Parkinson’s disease and dementia with Lewy bodies patients. Neurochem Int, 152, 105243 (2022).
24) Mahul-Mellier AL, Burtscher J, Maharjan N, Weerens L, Croisier M, Kuttler F, Leleu M, Knott GW, Lashuel HA. The process of Lewy body formation, rather than simply alpha-synuclein fibrillization, is one of the major drivers of neurodegeneration. Proc Natl Acad Sci USA, 117(9), 4971–4982 (2020).
25) Tsukada M, Ohsumi Y. Isolation and characterization of autophagy-defective mutants of Saccharomyces cerevisiae. FEBS Lett, 333(1-2), 169–174 (1993).

This page was created on 2022-06-22T11:32:25.523+09:00
This page was last modified on 2022-08-01T15:47:56.000+09:00
このサイトは(株)国際文献社によって運用されています。