全脳活動マップから紐解くストレス誘発不安応答の制御メカニズム
大阪大学 大学院薬学研究科神経薬理学分野 准教授
発行日:2022年12月30日Published: December 30, 2022
© 2022 日本神経化学会© 2022 The Japanese Society for Neurochemistry
急性ストレスは、不安などのネガティブな情動状態を出現させ、安全を得るための適切な行動選択を促す1, 2)。このストレス応答には、大脳皮質、皮質下、脳幹領域間のコミュニケーションによる神経細胞活動の大規模な変化が必要であり、それによりカテコールアミンシグナルの変化やそれに伴う行動反応が生じる1, 2)。過度や反復されるストレスによって、このストレス応答が破綻すると、不安障害やうつ病などの精神疾患が発症すると考えられている3)。したがって、ストレス応答のメカニズムの解明は、情動反応の制御という観点だけではなく、ストレス性精神疾患の病態発症の理解にも繋がると考えられる。そのため、これまでにもストレス応答の基盤となる神経ネットワークの全容解明に向け、関連する神経回路を同定する多くの研究が行われてきた4–7)。しかしながら、いまだに十分には解明されていないのが現状である。
その理由の一つに、未知の神経要素を同定するための仮説のないアプローチがほとんどなかったことが考えられる。特に、大規模なvolume imagingや機能的な細胞標識法に技術的な限界があった。この現状を打破するため、様々な全脳イメージング法が開発されてきた8, 9)。我々も、サブセルラーな空間解像度で脳全体を高速に撮影する全脳イメージングシステムFAST(block-face serial microscopy tomography)の開発に成功した10, 11)。そこで本稿では、FASTシステムと最初期遺伝子レポーターシステムを組み合わせて、これまで見落とされてきたストレス応答に関わる神経細胞集団および回路を同定した著者らの最近の研究成果12)を概説する。
社会的敗北ストレスや拘束ストレスは実験的な精神的ストレスモデルとして汎用されており、これらのストレスに曝されたマウスでは、不安関連行動などのストレス応答が表出される。このストレス応答を制御・調節するメカニズムにおいて、重要な細胞集団は、異なるストレスに対しても共通すると考えられる。そこで、社会的敗北ストレス、または拘束ストレスを負荷したマウスの脳と、ストレスに暴露しないホームケージ飼育のコントロール脳の活動状態を比較し、共通して変化する細胞集団を探索した。活性化した神経細胞の指標となる最初期遺伝子の一つArcのレポーター系であるArc-dVenusマウス13)にストレスを負荷し、FASTシステムを用いて全脳画像を撮影した。仮説フリーに重要な脳領域を明らかにするため、3次元再構成した脳画像を、形態学的な特徴に基づいて22の脳領域に分割し、各脳領域のdVenus陽性細胞を自動計数した。2つのストレス要因に反応する脳領域をデータ駆動型解析により特定するため、教師あり学習モデルであるサポートベクターマシーンを用いて、コントロール群とストレス群の線形判別分析を実施した。その結果、判別にかかる超平面関数への各脳領域の重み係数の絶対値の散布図から、社会的敗北ストレスまたは拘束ストレスのいずれかとホームケージコントロールの全脳活動マップの識別には、前障および扁桃体基底外側核が重要であることが明らかになった。本来、この線形サポートベクター判別分析の重み係数をそのまま判別の寄与度に解釈することは出来ないため14)、対応する活動パターンに変換したところ、前障および扁桃体基底外側核が共通して重要な活動であることが示された(図1)。また、高次元かつ低標本数のデータを用いた線形サポートベクター判別分析では、オーバーフィッティングの可能性に直面する。そこで、この結果の頑健性を高めるために、L1正則化(LASSO)による次元削減と変数選択を実施した。LASSO解析においても、ストレス脳とコントロール脳の判別にかかる変数領域として、前障が共通していた。
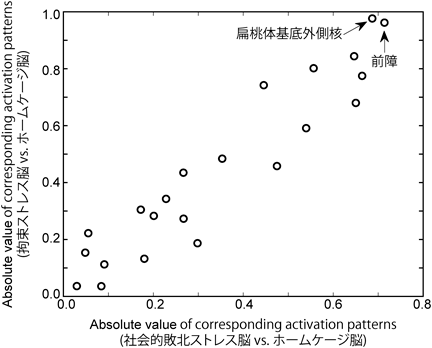
全22領域のdVenus陽性細胞数を計数し、ストレス脳(拘束ストレスまたは社会的敗北ストレス)とホームケージコントロール脳のサポートベクターマシーンによる判別分析を実施した(各n=5)。得られた判別分析の重みWを対応する生成モデルの活動パターンAをプロットした(A=∑xW∑ŝ−1; ∑は共分散、ŝは潜在要因)。
マウス脳の前障は吻尾側に伸びる長いシート状の構造をしている15, 16)。そこで前障の中で、ストレス暴露によって活性化するサブ領域を絞り込むため、前障におけるdVenus陽性神経細胞の空間分布を調べた。その結果、拘束ストレスおよび社会的敗北ストレスの両ストレス暴露後のdVenus陽性細胞の数は、前障の前方部分(ブレグマの前方約+1.52 mmから+0.96 mm)で共に有意に増加した。
次に、このストレス応答性前障神経細胞の機能的役割を明らかにするため、Targeted recombination in active populations(TRAP2)マウス17)とアデノ随伴ウイルス(AAV)を用いて、特異的な活動操作を行い行動表出への影響を調べた。マウスのストレス応答性前障細胞に興奮性DREADD受容体を発現させ特異的リガンドCNO投与により活性化させると、オープンフィールド試験において、中央ゾーンの滞在時間が有意に低下し、高架式十字迷路試験においても、オープンアーム滞在時間が有意に減少した(図2)。一方、マウスのストレス応答性前障細胞に抑制性DREADD受容体を発現させ、活動抑制するとストレス暴露により低下するオープンフィールド中の運動量および中央ゾーンの滞在割合が有意に増加した。
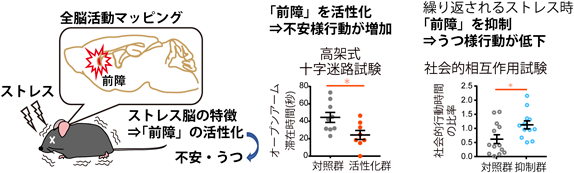
ストレス脳の判別分析により前障の活動の重要性を仮説フリーに同定した(図左)。この前障のストレス応答性神経細胞の活性化は高架式十字迷路のオープンアーム滞在時間を有意に低下させた(図中央)。また、反復ストレス時に前障ストレス応答性神経細胞の活動を抑制することにより、うつ様行動の表出が低下した。
ストレス暴露による不安関連行動は、脳内のカテコラミンシグナルの一過的な増加により惹起される1)。そこで、ストレス応答性前障細胞集団の活性化によって誘発される不安関連行動は、カテコラミンシグナルによって惹起されるのかを検討した。前障のストレス応答性神経細胞を活性化する30分前にドーパミンD2受容体拮抗薬ラクロプリドまたはβアドレナリン受容体拮抗薬プロプラノロールを投与すると、前障ストレス応答性神経細胞の活性化による不安関連行動が有意に抑制された。一方、ドーパミンD1受容体拮抗薬SCH39166またはα1-アドレナリン受容体拮抗薬プラゾシンの前処理は、不安関連行動に対して効果を示さなかった。これらの結果は、ストレス応答性前障神経細胞による不安関連行動の変化にカテコラミンシグナルが関与し、通常のストレス後の脳内反応を再現することを示唆している。
次に前障に神経投射し、ストレスによって活性化させる上流の細胞集団の脳内分布を調べるため、逆行性AAVとTRAP2マウスを用いてCre依存性の逆行性トレーシングを行った。その結果、特に扁桃体基底外側核にEGFP陽性神経細胞が多く存在することを見出した。これは、ストレスにより活性化した扁桃体基底外側核から前障に情報が伝達されていることを示している。
そこで次に、扁桃体基底外側核のストレス応答性神経細胞から前障への神経投射の活性化が不安関連行動を誘導するかについて、オプトジェネティクスを用いて調べた。TRAP2マウスの両側の扁桃体基底外側核にAAV-CaMKIIα-DIO-Chronos-EGFPを注入し、社会的敗北ストレス暴露直前にタモキシフェン投与しストレス応答性神経細胞にChronos-EGFPを発現させた。このマウスの両側の前障に光ファイバーを挿入し、リアルタイム場所嗜好性テストとオープンフィールドテストを実施した。リアルタイム場所嗜好性試験において、Chronos-EGFP発現マウスでは、前障に投射する扁桃体基底外側核のストレス応答性細胞の神経終末を光刺激しても運動量は変化しないものの、非光刺激区画に比べ光刺激区画での滞在時間が有意に短縮した。また、各3分間のOFF-ON-OFFの光刺激を行う9分間のオープンフィールド試験において、Chronos-EGFP発現マウスは、ベースラインのOFFエポックと比較してONエポック中の運動量に影響を与えずに、EGFP対照マウスと比較して光照射時に中央ゾーンでの滞在時間の減少を示した。これらのことから、扁桃体基底外側核のストレス神経細胞から前障に投射する神経回路は、ストレス誘発不安関連行動を惹起するのに十分であることが明らかになった。
最後に、ストレス応答に重要な前障の神経細胞集団がうつ様行動の発現にも関与するのかを検討した。うつ病モデルとして10日間連続した社会的敗北ストレスを負荷する慢性社会的敗北ストレスを用いた。TRAP2マウスのストレス応答性前障神経細胞に抑制性DREADD受容体hM4Di-mCherryを発現させ、各ストレス暴露の30分前にCNOを投与し活動を抑制したマウスは、社会的相互作用試験中の相互作用比率の低下が有意に抑制された。また、うつ症状の一つである無快感症の指標となるスクロース嗜好性も有意に増加した。これらの結果から、ストレス暴露時のストレス応答性前障神経細胞の活動抑制は、うつ発症を抑制できることが示された(図2)。
扁桃体基底外側核が負の情動行動を制御する統合的なハブであることはよく知られている5, 6, 18, 19)。扁桃体基底外側核からより大きな脳ネットワークへの負の情動シグナルの伝達は、状態依存的な感情状態や行動の調節に重要な役割を果たすと考えられるが、その下流にある経路はまだ解明されていなかった6)。本研究で行った仮説フリーなストレス脳の判別分析、機能的細胞標識、光刺激による回路操作により、前障は機能的にも解剖学的にも扁桃体基底外側核とつながっており、前障のストレス応答性神経細胞の集団が扁桃体基底外側核の下流で重要な機能的役割を果たしていることを初めて明らかにした。本稿では、全脳全細胞の活動状態から仮説フリーに解析し、これまで知られていなかった情動過程を調節する神経回路を同定した一例を紹介した。
脳機能をシステムレベルで理解することは、単なる器官機能の理解や医学の進歩にとどまらず、“心”の理解にも繋がる可能性を秘めた、人類の最大の課題の一つといえる。これを解決するためには、多様な細胞集団を詳細に理解することが必要不可欠である。上述の全脳神経活動の観察だけでなく、脳の半分を占めるとされるグリア細胞の形態変化も重要な課題である。実際、グリア細胞の一つであるアストロサイトの中枢神経系への機能や疾患への関与が示されつつある。過去15年間、様々な分野の研究ツールが飛躍的に進歩したため、ようやく脳の体系的な理解が進みつつある。今後のさらなる技術革新により、脳やこころの理解が益々深まることを期待したい。
本研究成果は、大阪大学大学院薬学研究科 神経薬理学分野にて得られたものです。研究遂行にあたり、多大なご指導を賜りました橋本均教授、共同研究者の先生方、ならびに実験を遂行してくれた研究室のメンバーに感謝いたします。また、本稿で紹介した研究内容は、日本学術振興会、文部科学省、日本医療研究開発機構、科学技術振興機構、武田科学振興財団、持田記念医学薬学振興財団、蓬庵社からの研究費により行われました。最後に、本稿の執筆機会を与えて下さいました日本神経化学会出版・広報委員会、ならびに優秀賞選考委員会の皆様にこの場を借りて御礼申し上げます。
1) Hermans EJ, Henckens MJA, Joëls M, Fernández G. Dynamic adaptation of large-scale brain networks in response to acute stressors. Trends Neurosci, 37(6), 304–314 (2014).
2) Lupien SJ, McEwen BS, Gunnar MR, Heim C. Effects of stress throughout the lifespan on the brain, behavior and cognition. Nat Rev Neurosci, 10(6), 434–445 (2009).
3) Gross C, Hen R. The developmental origins of anxiety. Nat Rev Neurosci, 5(7), 545–552 (2004).
4) McEwen BS, Nasca C, Gray JD. Stress effects on neuronal structure: Hippocampus, amygdala, and prefrontal cortex. Neuropsychopharmacology, 41(1), 3–23 (2016).
5) Tovote P, Fadok JP, Lüthi A. Neuronal circuits for fear and anxiety. Nat Rev Neurosci, 16(6), 317–331 (2015).
6) Gründemann J, Bitterman Y, Lu T, Krabbe S, Grewe BF, Schnitzer J, Lüthi A. Amygdala ensembles encode behavioral states. Science, 364(6437), eaav8736 (2019).
7) Calhoon GG, Tye KM. Resolving the neural circuits of anxiety. Nat Neurosci, 18(10), 1394–1404 (2015).
8) Osten P, Margrie TW. Mapping brain circuitry with a light microscope. Nat Methods, 10(6), 515–523 (2013).
9) Ueda HR, Ertürk A, Chung K, Gradinaru V, Chédotal A, Tomancak P, Keller PJ. Tissue clearing and its applications in neuroscience. Nat Rev Neurosci, 21(2), 61–79 (2020).
10) Seiriki K, Kasai A, Hashimoto T, Schulze W, Niu M, Yamaguchi S, Nakazawa T, Inoue KI, Takada M, Naka Y, Igarashi H, Tanuma M, Waschek JA, Ago Y, Tanaka KF, Hayata-Takano A, Nagayasu K, Shintani N, Hashimoto R, Kunii Y, Hino M, Matsumoto J, Yabe H, Nagai T, Fujita K, Matsuda T, Takuma K, Baba A, Hashimoto H. High-speed and scalable whole-brain imaging in rodents and primates. Neuron, 6(6), 1085–1100 (2017).
11) Seiriki K, Kasai A, Nakazawa T, Niu M, Naka Y, Tanuma M, Igarashi H, Yamaura K, Hayata-Takano A, Ago Y, Hashimoto H. Whole-brain block-face serial microscopy tomography at subcellular resolution using FAST. Nat Protoc, 14(5), 1509–1529 (2019).
12) Niu M, Kasai A, Tanuma M, Seiriki K, Igarashi H, Kuwaki T, Nagayasu K, Miyaji K, Ueno H, Tanabe W, Seo K, Yokoyama R, Ohkubo J, Ago Y, Hayashida M, Inoue KI, Takada M, Yamaguchi S, Nakazawa T, Kaneko S, Okuno H, Yamanaka A, Hashimoto H. Claustrum mediates bidirectional and reversible control of stress-induced anxiety responses. Sci Adv, 8(11), eabi6375 (2022).
13) Eguchi M, Yamaguchi S. In vivo and in vitro visualization of gene expression dynamics over extensive areas of the brain. Neuroimage, 44(4), 1274–1283 (2009).
14) Haufe S, Meinecke F, Görgen K, Dähne S, Haynes JD, Blankertz B, Bießmann F. On the interpretation of weight vectors of linear models in multivariate neuroimaging. Neuroimage, 87, 96–110 (2014).
15) Mathur BN. The claustrum in review. Front Syst Neurosci, 8, 48 (2014).
16) Goll Y, Atlan G, Citri A. Attention: the claustrum. Trends Neurosci, 38(8), 486–495 (2015).
17) DeNardo LA, Liu CD, Allen WE, Adams EL, Friedmann D, Fu L, Guenthner CJ, Tessier-Lavigne M, Luo L. Temporal evolution of cortical ensembles promoting remote memory retrieval. Nat Neurosci, 22(3), 460–469 (2019).
18) Tye KM, Prakash R, Kim SY, Fenno LE, Grosenick L, Zarabi H, Thompson KR, Gradinaru V, Ramakrishnan C, Deisseroth K. Amygdala circuitry mediating reversible and bidirectional control of anxiety. Nature, 471(7338), 358–362 (2011).
19) Felix-Ortiz AC, Beyeler A, Seo C, Leppla CA, Wildes CP, Tye KM. BLA to vHPC inputs modulate anxiety-related behaviors. Neuron, 79(4), 658–664 (2013).

This page was created on 2022-11-02T16:41:27.899+09:00
This page was last modified on 2023-01-11T14:04:20.000+09:00
このサイトは(株)国際文献社によって運用されています。