エピソード記憶の形成に必要な神経回路メカニズム
テキサス大学サウスウェスタン医学センター 精神医学部門 神経科学部門
発行日:2022年12月30日Published: December 30, 2022
© 2022 日本神経化学会© 2022 The Japanese Society for Neurochemistry
我々は、日々の生活の中でさまざまな出来事に遭遇し、それを覚える。これをエピソード記憶と呼び、「いつ」、「どこで」、「何が」、「誰と」という情報を含む1, 2)。エピソード記憶形成には大脳嗅内皮質–海馬回路が必要である3)。これまでの研究によって、「何が」、「どこで」の2つの情報がどのように脳の中で統合され、記憶されていくかについては比較的理解が進んでいる4)が、「いつ」という時間に関わる情報が、「何が」や「どこで」という情報と、どのように統合され、記憶されるのかについては明らかではなかった。「いつ」という時間情報において、特に、1)時間的に離れた2つの出来事を統合・分離すること、2)出来事間のタイミングを覚えること、3)出来事の順番を記憶することがエピソード記憶の形成に必須な機能として挙げられるが、それらを可能とする神経回路メカニズムは明らかではない。本研究では、「いつ」、「どこで」に関わる記憶情報の符号化に必要な神経細胞種を同定することを目的とした。
また、エピソード記憶の形成後、最初はその出来事を思い出すのに主に海馬を必要とするが、時間経過に伴い徐々に海馬は必要でなくなり、数週間後には大脳皮質を使ってその出来事を思い出す。この過程は記憶のシステムズコンソリデーションと呼ばれる5)。このことから、記憶は、時間経過とともに、海馬から大脳皮質に徐々に転送され、最終的には大脳皮質に貯蔵されると考えられている5)。しかし、この問題にアプローチする研究のほとんどは、いつ、どの脳部位が記憶の思い出しに“必要”なのかを調べる実験が主だったため、どの脳部位に記憶が“貯蔵”されているのか? そして、本当に海馬から大脳皮質へ記憶情報の転送は起きているのか? といった問題に対し、記憶を担う細胞を標識する方法論がなかったために、直接的に答えることが出来なかった。本研究では、記憶を担う細胞(記憶痕跡細胞またはエングラム細胞と呼ばれる)を標識する方法6)と光遺伝学7)を組み合わせることによって、記憶固定化の過程でどの脳部位に記憶エングラムが存在するのかを調べ、システムズコンソリデーションに必要な記憶エングラム細胞とその神経回路を同定することを目的とした。
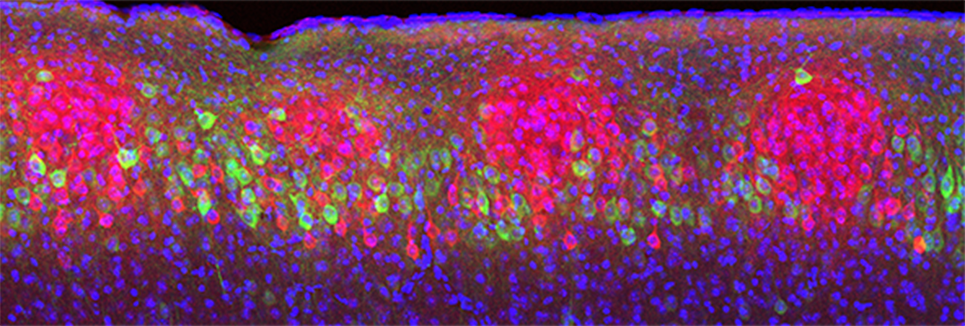
大脳嗅内皮質II層の神経細胞の多くは、海馬の歯状回に軸索を投射していることが知られていた8)。これを実証するために、海馬歯状回に逆行性トレーサーを注入し、大脳嗅内皮質II層のどの神経細胞が歯状回に投射しているのかを調べた。すると非常に面白いことに、II層内でトレーサーによって染色されない神経細胞群が存在し、それらの神経細胞は球状の細胞集団(クラスター)を形成していた(図1)9)。この細胞集団に特異的な遺伝子群を調べることにより、II層には歯状回に軸索を投射するReelin陽性の星形細胞と海馬CA1に投射するWfs1陽性の錐体細胞の2種類の興奮性神経細胞集団が存在することが分かった(図1)9)。Wfs1陽性細胞は100個程度の神経細胞からなる球状の細胞クラスターを形成し、約30個の細胞クラスターが格子状に大脳嗅内皮質II層に内包されていた(図1)9)。構造上の特徴から、このWfs1陽性細胞をアイランドセル、Reelin陽性細胞をオーシャンセルと名付けた9)。アイランドセルは海馬のCA1領域のSL層に軸索を投射し抑制性神経細胞とシナプスを作ることで、海馬CA1の神経活動に対して抑制的に働くことが分かった(図2)9)。過去の研究から、数秒から数十秒にわたって時間的に離れた2つの出来事を1つのエピソードとして記憶するためには、大脳嗅内皮質III層から海馬CA1への直接入力が必須ということが分かっていた10)。しかし、この機能は、動物の行動利潤のために、適切に調節される必要がある。例えば、時間的に離れすぎている2つの出来事、もしくは、印象が非常に弱い出来事同士などは覚える必要がなく、そうしないと、脳の中で記憶の混乱が生じる。しかし、そのような時間的に離れた2つの出来事の連結分離を調節するような仕組みは全く知られていなかった。我々は、アイランドセルが海馬の抑制性神経細胞を介して、大脳嗅内皮質III層の海馬への入力を抑制することから、アイランドセルが時間的に離れた2つの出来事を1つのエピソードとして記憶することを抑制するという仮説を立て検証した。その結果、アイランドセルを活性化すると、連結すべき2つの出来事が連結できなかった。逆に、アイランドセルを抑制すると本来連結すべきでない2つの出来事が1つのエピソードとして記憶された9)。一方オーシャンセルを抑制しても影響がなかった。以上のことから、大脳嗅内皮質III層から海馬CA1への神経入力がアクセルとして、アイランドセルから海馬CA1への入力がブレーキとして働き、時間的に離れた出来事の連結と分離が調節されることが分かった9)。本研究では、新規の神経回路を発見しただけでなく、時間的に離れた2つの出来事の連結と分離をアイランドセルが制御することを発見した。遺伝子発現にも正と負の調節機構が存在するが、今回、記憶形成にもそうした調節機構が神経回路レベルで存在することが初めて明らかになった。
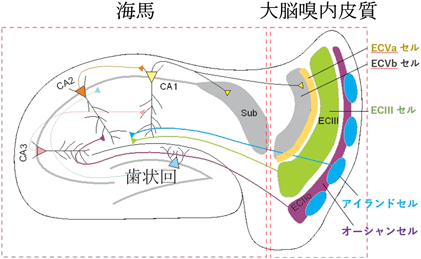
次に、大脳嗅内皮質II層のオーシャンセルはエピソード記憶の形成にどのような役割を果たすのかを検討した。オーシャンセルは海馬歯状回の顆粒細胞、CA3錐体細胞に投射しシナプスを形成する(図2)。これまでの研究から、海馬歯状回、CA3細胞は、環境の記憶形成に重要な役割を果たすと考えられてきた11–13)が、上流にあたるオーシャンセルが環境記憶に関してどのような役割を果たすのか不明であった。そこで、カルシウムに結合すると蛍光輝度が変化する蛍光タンパク質GCaMP614)を、オーシャンセルもしくはアイランドセルにそれぞれ細胞腫特異的に発現させ、オーシャンセル、アイランドセルの神経活動を超小型顕微鏡15)を用いて自由行動下で観察した。その結果、マウスが特定の部屋Aを探索すると、あるオーシャンセルは非常に強く神経活動を示すのに対して、マウスが違う部屋Bに入った途端、神経活動が弱くなる16)。逆に、部屋Bではほとんど活動しないが、部屋Bに入った途端、強い神経活動を示すオーシャンセルも観察された16)。アイランドセルでは、そのような環境特異的な神経活動を示す細胞は、観察されなかった。オーシャンセルの神経活動を特異的に抑制することによって、オーシャンセルが海馬CA3細胞の環境特異的神経活動を生成していること、マウスが異なる環境を区別して覚えるのにオーシャンセルが必須であることがわかった16)。一方、アイランドセルを抑制してもそれらに影響はなかった16)。更に海馬の神経活動を抑制しても、オーシャンセルで見られる環境特異的な神経活動は阻害されないこと16)から、オーシャンセルが環境情報を海馬に伝達していることが分かった。これまでは海馬(歯状回—CA3)において環境情報が生成されると考えられてきたが、この実験によって、海馬の上流にある大脳嗅内皮質II層のオーシャンセルが既に環境情報「どこで」をコードし、その情報が海馬に送られることによって、動物が周りの環境を区別して覚えることができることが分かった(図2)。
II層と同様に、大脳嗅内皮質V層でも異なる遺伝子マーカー、軸索投射パターンを持つ2種類の細胞集団がいることが分かった(図2)17)。Va細胞は、主に外に大脳嗅内皮質の外に軸索を伸ばし、Vb細胞は大脳嗅内皮質の中に軸索が留まる17)。Va細胞は大脳皮質の前頭前皮質に軸索を伸ばす17)。これまで記憶獲得後に海馬から前頭前皮質に記憶が転送されると考えられていた18)が、海馬と前頭前皮質をつなぐ神経回路が記憶の固定化に関与するかどうかは不明だった。そこで、Va細胞から前頭前皮質への神経投射を学習中に抑制すると、学習1日後、7日後の記憶想起テストではその影響は見られなかったが、14日後の再度の記憶テストで障害がみられた17)。このことから、学習中の海馬から前頭前皮質へ神経入力が14日後の記憶想起に必須であることがわかった。次に、記憶を担うエングラム細胞を標識する方法6)と光遺伝学7)を組み合わせることで、いつ前頭前皮質にエングラム細胞が形成されるのかを調べた。cfos-tTAシステム19)を用い、学習時に活性化した細胞にチャネルロドプシン7)を発現させ、その細胞を人工的に青色光で興奮させた。その結果、学習1日後に4 Hzの青色光で刺激すると、マウスはショックを受けた環境に入らなくても、恐怖の記憶を想起し、すくみ反応を示した17)。つまり、これまで考えられてきた記憶固定化の標準モデル5)とは異なり、学習1日後には既に前頭前皮質でエングラム細胞が形成されていた。また、学習時における大脳嗅内皮質Va細胞から前頭前皮質への神経入力は、前頭前皮質のエングラム細胞の生成に必須であった17)。さらに、学習時に前頭前皮質で生成されたエングラム細胞が実際の記憶想起に必要なのかを調べた。学習後1日では、前頭前皮質のエングラム細胞は実際の記憶想起に必要ではなかった。しかし時間の経過とともに、前頭前皮質のエングラム細胞で樹状突起の数が増大し、2週間後には、実際の記憶想起に必要となった17)。以上の結果から、前頭前皮質のエングラム細胞は、最初から記憶情報は持っているけれどもすぐには想起に使えない状態にある。しかし、時間経過とともに、前頭前野での神経細胞同士のつながりを強化し、2週間後には、実際の記憶想起に必要となる。これらの実験結果から、記憶は海馬から大脳皮質に転送されるのではなく、学習時に生成された前頭前野のエングラム細胞が時間経過とともに機能的に成熟することで大脳皮質での長期記憶が成立することがわかった17, 18)。
本研究は、分子マーカーを利用することによって、細胞種特異的な標識・操作を可能とし、マウス脳において新規神経細胞群を発見した。更に、その細胞群の記憶学習への役割や神経生理学的特徴をも同定した。これにより、これまでばらばらの情報として離散していた各神経細胞群の解剖学的情報、神経生理学的情報、脳機能情報が直接連結され、記憶学習に関わる脳神経回路を統合的に理解することが可能となった。アイランドセルが存在する大脳嗅内皮質はアルツハイマー疾患過程で、最初に影響を受ける脳領域の1つである。アルツハイマー患者の死後脳解析では大脳嗅内皮質II層でアイランドセルの細胞クラスターが消失していること20, 21)から、アイランドセルの研究は認知症を改善するための創薬ターゲットの可能性がある。また、ヒトの大脳嗅内皮質II層でのアイランドセルの細胞クラスターの数が、げっ歯類に比べ発達していること22)から、なぜアイランドセルがセルクラスターを形成するのかの疑問は、高次機能の進化の理解につながると考えられる。システムズコンソリデーションは、当時、記憶を物理化学的に測る方法論が存在しなかったことから、記憶がどの脳領域に存在するのかを同定できなかったため、対立する仮説は決着がつかず、システムレベルでの記憶固定化の仕組みについて膠着状態にあった。本研究は、記憶エングラム細胞が海馬と大脳皮質でいつ形成され消去されるかを検証することで、どちらの考えも部分的に合っているが部分的に間違っていることもわかり、結果、新しい記憶固定化のメカニズムを提唱することができた17, 18)。
本研究は、マサチューセッツ工科大学の利根川進先生の指揮のもと実施されました。利根川先生はもちろん、本研究に協力していただきました多くの共同研究者に深く感謝いたします。2022年度日本神経化学会優秀賞に選んでいただきまして誠にありがとうございました。
1) Marr D. Simple memory: a theory for archicortex. Philos Trans R Soc Lond B Biol Sci, 262(841), 23–81 (1971).
2) Tulving E. Episodic memory: from mind to brain. Annu Rev Psychol, 53(1), 1–25 (2002).
3) Eichenbaum H. A cortical-hippocampal system for declarative memory. Nat Rev Neurosci, 1(1), 41–50 (2000).
4) Knierim JJ, Neunuebel JP, Deshmukh SS. Functional correlates of the lateral and medial entorhinal cortex: objects, path integration and local-global reference frames. Philos Trans R Soc Lond B Biol Sci, 369(1635), 20130369 (2014).
5) Squire LR. Mechanisms of memory. Science, 232(4758), 1612–1619 (1986).
6) Liu X, Ramirez S, Pang PT, Puryear CB, Govindarajan A, Deisseroth K, Tonegawa S. Optogenetic stimulation of a hippocampal engram activates fear memory recall. Nature, 484(7394), 381–385 (2012).
7) Yizhar O, Fenno LE, Davidson TJ, Mogri M, Deisseroth K. Optogenetics in neural systems. Neuron, 71(1), 9–34 (2011).
8) Tamamaki N, Nojyo Y. Projection of the entorhinal layer II neurons in the rat as revealed by intracellular pressure-injection of neurobiotin. Hippocampus, 3(4), 471–480 (1993).
9) Kitamura T, Pignatelli M, Suh J, Kohara K, Yoshiki A, Abe K, Tonegawa S. Island cells control temporal association memory. Science, 343(6173), 896–901 (2014).
10) Suh J, Rivest AJ, Nakashiba T, Tominaga T, Tonegawa S. Entorhinal cortex layer III input to the hippocampus is crucial for temporal association memory. Science, 334(6061), 1415–1420 (2011).
11) Leutgeb JK, Leutgeb S, Moser MB, Moser EI. Pattern separation in the dentate gyrus and CA3 of the hippocampus. Science, 315(5814), 961–966 (2007).
12) McHugh TJ, Jones MW, Quinn JJ, Balthasar N, Coppari R, Elmquist JK, Lowell BB, Fanselow MS, Wilson MA, Tonegawa S. Dentate gyrus NMDA receptors mediate rapid pattern separation in the hippocampal network. Science, 317(5834), 94–99 (2007).
13) Nakashiba T, Young JZ, McHugh TJ, Buhl DL, Tonegawa S. Transgenic inhibition of synaptic transmission reveals role of CA3 output in hippocampal learning. Science, 319(5867), 1260–1264 (2008).
14) Chen TW, Wardill TJ, Sun Y, Pulver SR, Renninger SL, Baohan A, Schreiter ER, Kerr RA, Orger MB, Jayaraman V, Looger LL, Svoboda K, Kim DS. Ultrasensitive fluorescent proteins for imaging neuronal activity. Nature, 499(7458), 295–300 (2013).
15) Ziv Y, Burns LD, Cocker ED, Hamel EO, Ghosh KK, Kitch LJ, El Gamal A, Schnitzer MJ. Long-term dynamics of CA1 hippocampal place codes. Nat Neurosci, 16(3), 264–266 (2013).
16) Kitamura T, Sun C, Martin J, Kitch LJ, Schnitzer MJ, Tonegawa S. Entorhinal cortical ocean cells encode specific contexts and drive context-specific fear memory. Neuron, 87(6), 1317–1331 (2015).
17) Kitamura T, Ogawa SK, Roy DS, Okuyama T, Morrissey MD, Smith LM, Redondo RL, Tonegawa S. Engrams and circuits crucial for systems consolidation of a memory. Science, 356(6333), 73–78 (2017).
18) Tonegawa S, Morrissey MD, Kitamura T. The role of engram cells in the systems consolidation of memory. Nat Rev Neurosci, 19(8), 485–498 (2018).
19) Reijmers LG, Perkins BL, Matsuo N, Mayford M. Localization of a stable neural correlate of associative memory. Science, 317(5842), 1230–1233 (2007).
20) Gomez-Isla T, Price JL, McKeel DW Jr., Morris JC, Growdon JH, Hyman BT. Profound loss of layer II entorhinal cortex neurons occurs in very mild Alzheimer’s disease. J Neurosci, 16(14), 4491–4500 (1996).
21) Mikkonen M, Alafuzoff I, Tapiola T, Soininen H, Miettinen R. Subfield- and layer-specific changes in parvalbumin, calretinin and calbindin-D28K immunoreactivity in the entorhinal cortex in Alzheimer’s disease. Neuroscience, 92(2), 515–532 (1999).
22) Naumann RK, Ray S, Prokop S, Las L, Heppner FL, Brecht M. Conserved size and periodicity of pyramidal patches in layer 2 of medial/caudal entorhinal cortex. J Comp Neurol, 524(4), 783–806 (2016).

This page was created on 2022-11-02T18:36:44.312+09:00
This page was last modified on 2023-01-19T15:53:59.000+09:00
このサイトは(株)国際文献社によって運用されています。