ヒトのニューロンにおける神経突起伸長メカニズムの解析
慶應義塾大学医学部生理学教室
発行日:2022年12月30日Published: December 30, 2022
© 2022 日本神経化学会© 2022 The Japanese Society for Neurochemistry
私が所属する慶應義塾大学医学部生理学教室では、これまでに亜急性期脊髄損傷に対するヒトiPS細胞由来神経幹細胞/前駆細胞(human induced pluripotent stem cell derived neural stem/progenitor cell; hiPSC-NS/PC)移植による再生治療研究を行ってきており、当院整形外科学教室とともに、2021年末から世界初の臨床研究も開始されている。しかし、脊髄損傷患者の大部分は慢性期の患者さんであり、かつ、慢性期脊髄損傷の治療は、亜急性期脊髄損傷の治療に比べ、損傷部の環境の変化が軸索再生に不利に働くなど、クリアすべき課題がいくつか存在しており、慢性期脊髄損傷の治療技術の開発が求められているのが現状である。
また、我々は、これまでにγ-セクレターゼ阻害剤(γ-secretase inhibitor; GSI)のDAPTをhiPSC-NS/PCに添加した後に、hiPSC-NS/PCを損傷患部へ移植することで、分化したニューロンの神経突起が伸長し慢性期脊髄損傷モデルマウスを治療することに成功してきた1)。
しかし、ヒトのニューロンにおいて、神経突起伸長がどのようなメカニズムで生じているのか、どのようにすれば伸長を促進できるか解明できていない点があり、その解明を目的に本研究を開始した。神経突起の伸長メカニズムの研究は、これまでヒト以外のニューロン(マウス、ラット、ゼブラフィッシュなど)で行われてきたが、本研究は、今後の慢性期脊髄損傷の再生治療への応用を見据えて、hiPSCから作製したヒトのニューロンに着目して行った。
最初に、神経突起伸長を促進するシグナルを同定するためのスクリーニング実験を行った。hiPSC 201B72)、およびhiPSC 414C23)の2ラインからhiPSC-NS/PC(neurosphere)を誘導し、GSIであるDAPTとCompound 34をこれらneurosphereに添加して神経系の遺伝子発現を促進させた。Control群、DAPT添加群、Compound 34添加群の各々の群について直径100 µmのneurosphereを選択し、GSI(DAPT、compound 34)により神経系の遺伝子を発現させたこれらサンプルをスクリーニングサンプルとして、RNA-seqにより全転写産物を解析した。ちなみに、γ-セクレターゼ阻害剤の選択に関しては、予備実験にてDAPTとCompound 34がhiPSC-NS/PCにおいて最もγ-セクレターゼ阻害作用が強く認められた事に加え、単一の化合物特有の効果ということを排除するために、これら2つを選択した。
我々の以前の報告において、hiPSC-NS/PCをDAPTで処理すると、mitogen-activated protein kinase(MAPK)シグナルが増強し、神経突起伸長が促進されることが示唆されていた1)。そこで、DAPTまたはCompound 34の投与後に発現が変動した全遺伝子のうち、MAPKに関連する遺伝子を選択し、さらに解析した。その結果、MAPKの中で、growth arrest and DNA damage gamma(GADD45G)の発現が最も大きく上昇し、GADD45GはMAPK関連遺伝子全体の代表として適していることを数学的解析から明らかにした。さらに、GSIの投与により発現が変化した全遺伝子の中でも、GADD45Gは最も高い発現を示したひとつであった。
さらに、MAPK関連遺伝子の発現レベルおよび、MAPK関連タンパク質のリン酸化状態を免疫染色、ウエスタンブロッティングにて調べていくと、p38とCDC25Bのリン酸化レベルがGSI投与群で有意に上昇していたため、GSI処理後に発現変化を示すGADD45、p38、CDC25Bが同じシグナル伝達経路で作用しているかどうかを検証するために複数の検証実験を行った。
まず、GADD45GのsiRNAによるノックダウンを行うと、neurosphereをGSIで処理しても、リン酸化p38レベルが有意に減少したことから、GADD45Gがp38の上流で機能していることが明らかとなった。次に、リン酸化CDC25Bのレベルは、GSI処理にもかかわらず、SB203580(p38 MAPK阻害剤)により抑制された。これら実験により、これらは同一のシグナル伝達経路内で作用し、GADD45G→p38のリン酸化→CDC25Bのリン酸化の順でシグナル経路が伝達されることが明らかになった。
また、Neurosphereにおいて、レンチウイルスベクターを用いてGADD45Gを過剰発現させると、p38の総発現量はcontrol群とGADD45G過剰発現群の間で差がなかったが、p38のリン酸化レベルはGADD45G過剰発現で有意に上昇した。
将来の臨床応用を見据えて、化合物によるこのシグナル経路の活性化にも挑戦した。Streptomyces属から単離される化合物RK-682は、p38のチロシン残基の脱リン酸化を防ぐことができ4)、実際、本研究で用いたneurosphereの実験系でもp38の活性を維持することにつながり、RK-682処理後にリン酸化p38およびリン酸化CDC25Bのレベルが有意に上昇することが確認された。ちなみにRK-682は直接CDC25Bのリン酸化促進には働かないことが知られており4)、その面からもp38の下流にCDC25Bが位置していることが考えられた。
次に、このシグナル伝達経路の活性化が、実際に神経突起伸長につながるかどうかを検証した。これらレンチウイルスを用いた遺伝子導入および、RK-682添加のどちらでシグナル経路を増幅させた場合でも、neurosphereからニューロンに分化させると、微小管が伸長していることが微小管構成タンパク質であるα-tubulinとβ-III tubulinの免疫染色により明らかになり、タウの免疫染色によって神経突起が伸長していることが確認された。
次に、このシグナル経路が最終的に何に作用して微小管伸長、神経突起伸長を生じさせているのかを調べた。ここで、コラプシン反応媒介タンパク質2(collapsin response mediator protein 2; CRMP2)に着目した。CRMP2はトレオニンの514番目残基(Thr514)のリン酸化が脱リン酸化されると、tubulinに作用して微小管のアセンブリが進行して神経突起を伸ばすことが知られている5)。neurosphere由来のニューロンへのRK-682の添加後に、リン酸化CRMP2の量をウェスタンブロッティングにより定量したところ、そのリン酸化レベルが減少していることがわかった。
この解析から、RK-682投与群における神経突起伸長は、CRMP2の脱リン酸化により、微小管の重合が促進されることにより生じることが明らかになった。
また、GADD45G/p38 MAPK/CDC25Bシグナル経路の各シグナルをブロックすることで、神経突起伸長の表現型がキャンセルされるかどうかも検証した。neurosphereをSB203580(p38シグナル阻害剤)およびGSIとともに14日間培養してニューロン分化を誘導した後、β-III tubulinとタウの免疫染色を行ったところ、神経突起伸長はSB203580によるp38シグナルの阻害により抑制された。
さらに、下流エフェクターであるCDC25BをsiRNAでノックダウンした場合の効果についても検証した。CDC25Bに対するsiRNAとGSI双方をneuroshereに添加し、14日間培養するとニューロンへ分化するものの、CDC25Bのノックダウンにより、GSI処理にもかかわらず神経突起伸長が抑制された。CDC25Bをノックダウンしても、ニューロンへの分化は抑制されなかったことから、CDC25Bのシグナルはニューロンへの分化に影響を与えることなく神経突起伸長を誘導していることが示唆された。また、CRMP2のThr514のリン酸化の程度は、CDC25Bに対するsiRNA処理をした群の方が、対照群よりも高いことが分かり、CRMP2のリン酸化状態はCDC25Bシグナルにより減弱することで、神経突起伸長を促進することが示された。以上のことから、リン酸基を付加するGADD45Gを起点としたキナーゼカスケードが、CRMP2による微小管重合と神経突起伸長を促進することが明らかとなった。
また、神経突起の伸長には微小管の安定化も考慮に入れる必要があるため、微小管の安定化の指標となるα-tubulinのアセチル化(Lys40)を定量することで、このシグナル伝達経路が微小管の安定化に寄与しているかどうかを評価したが、アセチル化の状態は本シグナル経路とは関係がなかった。これらのデータを総合すると、本シグナル経路の神経突起伸長効果は、微小管の安定性よりも、微小管の重合反応の促進によってもたらされていることが示された。
ここまでに上述した実験では、hiPS-NS/PCをニューロンに分化させる際にGADD45Gの過剰発現や、RK-682による処理を施していたが、ニューロンに分化させた後にRK-682を加え、シグナル経路を増幅することでも、同様の神経突起伸長が認められるかも検証した。シングルセル状態のhiPSC-NS/PCをニューロンに分化させ、5日後にRK-682を投与し、RK-682によって引き起こされる神経突起伸長の程度を48時間のタイムラプスイメージングによって観察したところ、RK-682添加後に神経突起伸長が有意に増加した。したがって、細胞の状態がhiPS-NS/PCの幹細胞の状態でも、分化後のニューロンでも本シグナル経路は神経突起伸長に働くことが明らかになった。
最後に、ヒト脳におけるGADD45Gの発現をin vivoレベルで評価した。BrainSpan Atlas of the Developing Human Brain(http://www.brainspan.org)からヒト脳サンプルのバルクRNA-seqデータを取得し、胎児期から成人期までのヒト脳におけるGADD45Gの発現を解析した。GADD45Gの発現は、胎児の脳や10歳までのヒトの脳では、それ以上の年齢のヒトの脳よりも有意に高かった。さらに、GADD45Gの発現は、胎児期によっても異なり、初期胚期では後期胚期よりも高い傾向が認められた。このGADD45Gの高発現期には、ニューロンマーカーであるTUBBとDCXが高発現していたが、アストロサイトマーカーのSLC1A2とGLULは著しく発現量が減少していた。神経新生が活発な初期胚の時期にGADD45Gが高発現していることから、GADD45Gが神経突起伸長を含むニューロンの機能・形態面に関与している可能性が示唆された。
本研究では、hiPSC-NS/PC由来のニューロンにおいてGADD45Gから始まるシグナル経路が神経突起伸長を促進することを明らかにし、また、発達中のヒト胎児脳サンプルにおいても神経新生が盛んな時期と一致して、GADD45Gの発現が上昇していることを明らかにした(図1)。また、別の公開データベースでも、胚性幹細胞(ES細胞)由来の未熟なニューロンの分化・成熟に伴いGADD45Gの発現が上昇することが確認されており6)、本シグナル経路は胎児期の初期におけるニューロンの発達に伴い活性化される可能性が示唆された。
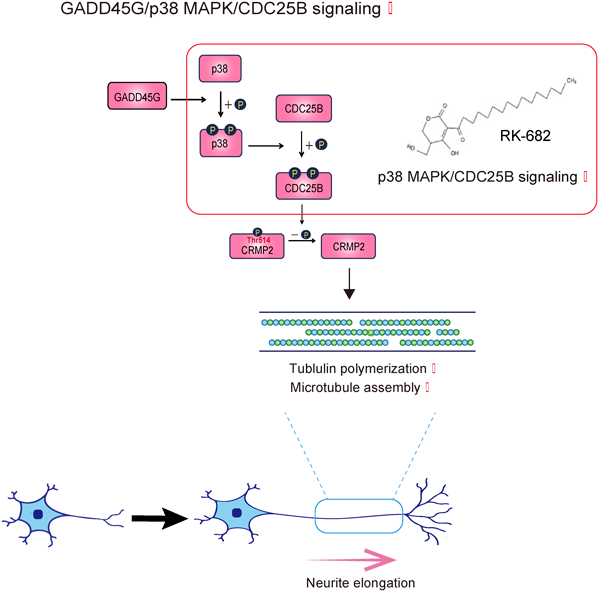
GADD45Gは、チンパンジーを含む非ヒト哺乳類で保存されているエンハンサー配列が、ヒトゲノムでは特異的に欠失されているhuman-specific conserved deletion enhancer-sequence(hCONDEL)として知られている7)。この欠失により、ヒト脳内のNS/PCではGADD45Gの発現が低下し、より増殖性の高い形質となり、それが進化の過程でヒトの脳の拡大・発達につながったと推測されている。本研究はhCONDELとして知られているGADD45Gのpost mitoticなニューロンでの役割を初めて解明したものであり、GADD45Gの精細な発現制御がヒトのニューロンの神経突起伸長を含む機能発揮に寄与している可能性があると考えられた。この発見は従前多く行われてきたヒト以外のマウスやゼブラフッシュなどを用いた解析では到達できなかったものであり、「ヒト」の細胞であるhiPSCを用いたから可能となったものである。
また、in vitroとin vivoの両方で認められる神経細胞系列でのGADD45Gの精細な発現上昇が、ヒトの成熟ニューロンにおける、幹細胞・前駆細胞の増殖状態から神経突起伸長状態への切り替えに重要な役割を果たすという仮説も考えられる。本研究結果も、この仮説を支持するものであり、GSIは増殖性の神経前駆細胞を、post mitoticなニューロンへと分化させるが1)、GSI処理したneurosphereにおけるGADD45Gの発現上昇は、神経突起伸長に重要な役割を果たすと考えられる。
本研究では、ヒト以外の動物種ではなく、ヒトのニューロンでの新たな神経突起伸長メカニズムを解明し、そのメカニズムに立脚した神経突起伸長化合物RK-682を同定したものであり、いまだ課題の残る慢性期脊髄損傷治療のブレイクスルーとなる可能性がある。
さらに、GADD45Gは、ヒトの大脳がなぜ他の動物種と異なり、大きく進化できたのかを説明できる遺伝子として注目されており、その遺伝子のニューロンにおける役割を初めて発見できた本研究結果は進化生物学的にも意義のあると考えられる。
慢性期脊髄損傷治療への応用もさる事ながら、GADD45Gの発現様式が異なるマウスとヒトでの神経突起伸長のメカニズムの差異の検証や、今回発見したシグナル経路の強度が老化とともに減少してしまうのかどうか、もしそうであるならば、このシグナル経路を増強すれば若返りにつながるのか、また他の神経軸索が変性してしまう神経変性疾患でも共通のメカニズムがあるのかどうかなど、今後の研究の広がりの可能性を感じさせる成果であると考えられる。
201B7と414C2ヒトiPS細胞株を提供してくださった京都大学iPS細胞研究所山中伸弥教授に感謝いたします。理化学研究所の上口裕之先生には本研究についてのご助言をいただき、FIRSTプログラムのプロジェクトアドバイザーである五條堀孝先生と長谷川真理子先生には、hCONDELを含むヒト特異的遺伝子とその役割について、貴重なアドバイスをいただきました。重ねて感謝申し上げます。
1) Okubo T, Iwanami A, Kohyama J, Itakura G, Kawabata S, Nishiyama Y, Sugai K, Ozaki M, Iida T, Matsubayashi K, Matsumoto M, Nakamura M, Okano H. Pretreatment with a γ-secretase inhibitor prevents tumor-like overgrowth in human iPSCderived transplants for spinal cord injury. Stem Cell Reports, 7(4), 649–663 (2016).
2) Takahashi K, Tanabe K, Ohnuki M, Narita M, Ichisaka T, Tomoda K, Yamanaka S. Induction of pluripotent stem cells from adult human fibroblasts by defined factors. Cell, 131(5), 861–872 (2007).
3) Okita K, Matsumura Y, Sato Y, Okada A, Morizane A, Okamoto S, Hong H, Nakagawa M, Tanabe K, Tezuka KI, Shibata T, Kunisada T, Takahashi M, Takahashi J, Saji H, Yamanaka S. A more efficient method to generate integration-free human iPS cells. Nat Methods, 8(5), 409–412 (2011).
4) Hamaguchi T, Sudo T, Osada H. RK-682, a potent inhibitor of tyrosine phosphatase, arrested the mammalian cell cycle progression at G1phase. FEBS Lett, 372(1), 54–58 (1995).
5) Yoshimura T, Kawano Y, Arimura N, Kawabata S, Kikuchi A, Kaibuchi K. GSK-3beta regulates phosphorylation of CRMP-2 and neuronal polarity. Cell, 120(1), 137–149 (2005).
6) van de Leemput J, Boles NC, Kiehl TR, Corneo B, Lederman P, Menon V, Lee C, Martinez RA, Levi BP, Thompson CL, Yao S, Kaykas A, Temple S, Fasano CA. CORTECON: a temporal transcriptome analysis of in vitro human cerebral cortex development from human embryonic stem cells. Neuron, 83(1), 51–68 (2014).
7) McLean CY, Reno PL, Pollen AA, Bassan AI, Capellini TD, Guenther C, Indjeian VB, Lim X, Menke DB, Schaar BT, Wenger AM, Bejerano G, Kingsley DM. Human-specific loss of regulatory DNA and the evolution of human-specific traits. Nature, 471(7337), 216–219 (2011).

This page was created on 2022-11-02T18:20:36.776+09:00
This page was last modified on 2023-01-11T14:00:53.000+09:00
このサイトは(株)国際文献社によって運用されています。