脱髄研究における新たな病態モデルマウスと病変標識法の開発
自治医科大学医学部解剖学講座組織学部門
発行日:2022年12月30日Published: December 30, 2022
© 2022 日本神経化学会© 2022 The Japanese Society for Neurochemistry
髄鞘(ミエリン)は脂質二重膜が何層にも重なった膜構造物であり、絶縁体として軸索の跳躍伝導を可能にするとともに、軸索との相互作用によって様々な神経活動に関与している。中枢ミエリンが障害される代表的な神経疾患に多発性硬化症(MS)、末梢ミエリンが障害される疾患にはギランバレー症候群や慢性炎症性脱髄性多発神経炎などがあげられる1–4)。これらの中枢および末梢神経系の難治性脱髄性疾患は脱髄による運動麻痺や感覚障害を主症状とし、発症原因が不明な点も多く、根治療は困難であるため特定疾患に分類されている。その中でもMSの患者数は本邦でも年々増加しており、全世界で280万人にも上る。現在MSの再発予防には免疫抑制薬が使用されているが、再発と寛解を繰り返す中で進行型へと移行し、最終的には自立歩行も困難となる。しかしながら、これらの脱髄性疾患に対して再ミエリン化を促進する治療法は開発されていない。特に、中枢脱髄に伴うオリゴデンドロサイトの機能低下によって引き起こされる再ミエリン化障害や二次性軸索変性は後遺症として問題となるため、これらを予防する治療法開発が重要である。
脱髄を伴う神経疾患の治療法開発には、脱髄モデルマウスによる再ミエリン化の評価が行われている5, 6)。しかしながら、国内外で頻繁に使用されている実験的自己免疫性脳脊髄炎(EAE)モデルやクプリゾンモデルは、脱髄範囲や神経症状の調節も困難で扱いにくい場合が多く、作製に時間と手間がかかるという問題点が挙げられた。また、脱髄による運動機能障害と再ミエリン化による運動機能回復は見られない場合もあり、経時的な脱髄と組織再生という、MS患者で見られる特徴的な臨床所見とは異なる点もある。
また、実験動物を用いた脱髄モデルでは局所的な脱髄部位を解析する必要がある。局所的な脱髄を誘発するには界面活性剤であるリゾレシチンを白質や坐骨神経に直接注入することで脱髄を誘発するモデルが存在する7, 8)。しかし、脱髄部位を確認するには組織を固定し、組織切片を作製して観察する必要がある。そのため、生化学的な解析による分子機構解析や脱髄および再生を電顕観察により評価を行うには脱髄部位のみを摘出する必要があるが、未標識の脱髄部位のみを摘出することは困難である。
本稿ではこれらの問題点を解決するために、筆者が開発したニュートラルレッド色素による脱髄病変標識法と内包脱髄モデルマウスについて概説する。
急性の脱髄早期には、活性化したミクログリアやマクロファージ、アストロサイトが病変部位に集積することが知られている。そこで、筆者は生染色にも使用される可溶化色素であり、これまでにマクロファージのリソソームに取り込まれることが知られているニュートラルレッドに着目した9, 10)。そして、脱髄マウスにニュートラルレッドを投与することで生体内の脱髄部位が標識され、肉眼的に病変領域が観察できるのではないかと仮説を立てた。
まず初めに脱髄誘導剤であるリゾレシチンを脳梁、脊髄、坐骨神経に直接注入し、中枢および末梢神経系の局所脱髄モデルマウスを作製した。次に脱髄のピークが観察されるリゾレシチン投与5–7日後の脱髄マウスから組織を摘出する2時間前にニュートラルレッドを腹腔内に投与した。その結果、脱髄マウスにニュートラルレッドを腹腔内投与するだけで脱髄部位を肉眼で観察できることを発見した11, 12)。これらのニュートラルレッドによる標識は再ミエリン化と共に減少し、再ミエリン化後のマウスではほとんど検出されなかった。次にニュートラルレッドの標識細胞を調べるために各種マーカーとの免疫組織染色を行ったところ、中枢の脱髄部位では活性化ミクログリアと炎症応答性アストロサイト、末梢の脱髄部位では活性化マクロファージや貪食中のシュワン細胞が標識され、特に細胞内のリソソームにニュートラルレッドが局在していることが明らかになった。最後にニュートラルレッドによる標識法が生化学的解析や電子顕微鏡観察に応用できるかを調べるために、ニュートラルレッドで標識された病変部位のみを摘出し、ウエスタンブロット解析や定量PCR法、フローサイトメトリー、電子顕微鏡観察に適用可能であることを明らかにした11, 12)。
この手法を用いることで、中枢および末梢神経組織中の脱髄部位を肉眼的な観察で同定し、病変組織のみを摘出して解析できるようになった(図1)。
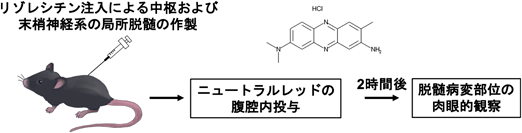
脳、脊髄、坐骨神経に直接リゾレシチンを注入した脱髄マウスにニュートラルレッドを腹腔内投与することで、脱髄部位のみを標識し、肉眼的に病変を観察することができる。
次に、リゾレシチンによる局所脱髄モデルと強力な血管収縮剤であるエンドセリン(ET1)による脳白質梗塞モデルを応用し7, 13)、新たなモデルマウスの開発に取り組んだ。皮質脊髄路は大脳皮質から内包白質を経由し、脊髄まで下降する経路であり、四肢の運動機能を制御している。また、これまでに多発性硬化症や白質梗塞により内包が障害されることで半身麻痺様の運動機能障害がみられることが報告されている14–17)。そこで、皮質脊髄路の主要経路である内包白質にリゾレシチンを局所的に注入し、脱髄させることで組織再生に加えて運動機能の回復が評価できる新たなモデルマウスになると仮説を立てた。内包脱髄マウスを作製するために、脳定位固定装置を用いてリゾレシチンを成熟マウスの内包に投与した。その結果、リゾレシチン投与7日後には内包の局所脱髄による半身麻痺様の運動機能障害が引き起こされ、運動機能障害が認められたマウスにニュートラルレッドを投与したところ内包に大きな脱髄が肉眼的に観察された。その後、ET1の内包内投与による脳白質梗塞モデルとは異なり、徐々に運動機能が回復し、リゾレシチン投与28日後にはPBS投与群と同レベルまで運動機能が回復することが明らかになった。さらに、内包脱髄マウスの詳細な組織解析を行ったところ、運動機能障害が見られるリゾレシチン投与7日後には大きな脱髄や炎症、軸索障害が観察されるのに対し、投与28日後には運動機能の回復と共に再ミエリン化が誘導され、炎症および軸索障害の軽減が見られることが明らかになった。したがって、本研究により樹立された内包脱髄モデルマウスは運動機能障害と再ミエリン化に伴う機能回復の両者が評価可能となる新たな脱髄モデルマウスになることが期待される18)(図2)。
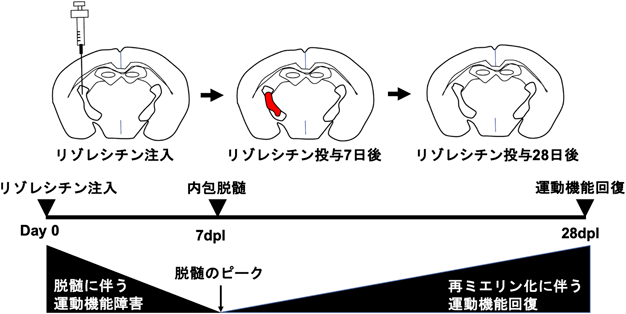
内包へのリゾレシチン注入により、投与7日後(7 dpl)には半身麻痺を伴う運動機能障害が誘導される。その後、投与28日後(28 dpl)まで再ミエリン化と共に運動機能の回復が見られる。
従来の脱髄研究では、脱髄と再ミエリン化およびそれに関連するシグナル経路の探索をするために、脱髄部位からサンプルの採取が行われてきた。しかしながら、脱髄および再ミエリン化部位を組織から選択的に採取するのは困難であり、サンプリングの際に含まれる非脱髄部位がノイズとして解析結果に影響することが十分に考えられた。また、脱髄および再ミエリン化の解析には電子顕微鏡による微細構造解析が不可欠であるが、組織を切り出して脱髄部位を探索するには経験と技術が必要なだけでなく、時間と手間がかかっていた。本稿で紹介した新たな病変標識法では、ニュートラルレッドを腹腔内投与するという非常にシンプルな方法で脱髄部位の肉眼観察を可能にさせ、これまでの問題点を解決することができた。
また、既存の脱髄モデルマウスでは再ミエリン化に伴う運動機能回復の評価は困難であった。そのため、基礎研究レベルで運動機能障害の回復を評価し、細胞保護や再ミエリン化の促進などの薬剤による治療効果を比較して、創薬につなげるための新たな動物モデルの開発が必要であった。内包脱髄モデルはリゾレシチンを内包内に投与するシンプルなモデルであることから、投与方法の調節によって脱髄範囲や神経症状の程度が調節可能である。したがって、系統によって感受性が変化するEAEモデルやクプリゾンモデルよりも扱いやすく、簡便かつ短期間に作製できるため、汎用性の高いモデルマウスであると確信している。
今後は、本研究を中枢および末梢神経系の再ミエリン化障害の原因究明およびMSや虚血性白質障害、末梢神経の脱髄を伴う難病に対する新たな治療法開発に展開したい。
日本神経化学会に入会してから、ちょうど10年という節目に大変名誉ある日本神経化学会奨励賞を賜り、光栄に思います。初めて学会に参加した時から、若手研究者育成セミナーには毎年参加させていただき、シニアの先生方だけでなく、多くの若手研究者と知り合う機会を与えてくださった日本神経化学会に心より感謝申し上げます。
本稿で紹介いたしました研究成果は、ジョージタウン大学生物学部と自治医科大学医学部解剖学講座組織学部門で得られました。多大なるご指導を賜りましたJeffrey Huang教授と大野伸彦教授に厚く御礼申し上げます。また、研究者として一から育てていただいた東京薬科大学馬場広子名誉教授(現新潟医療福祉大学教授)、山口宜秀准教授をはじめこれまでご指導いただきました諸先生方、本稿の執筆機会を与えて下さいました日本神経化学会優秀賞・奨励賞選考委員の先生方、並びに関係者の先生方に深く感謝申し上げます。今後も日本神経化学会奨励賞受賞者としての自覚を持ち、精進して研究に取り組みたいと考えております。今後ともご指導ご鞭撻のほど、よろしくお願い申し上げます。
1) Compston A, Coles A. Multiple sclerosis. Lancet, 372(9648), 1502–1517 (2008).
2) Dutta R, Trapp BD. Relapsing and progressive forms of multiple sclerosis: insights from pathology. Curr Opin Neurol, 27(3), 271–278 (2014).
3) Fadia M, Shroff S, Simpson E. Immune-mediated neuropathies. Curr Treat Options Neurol, 21(6), 28 (2019).
4) Yuki N, Hartung HP. Guillain-Barre syndrome. N Engl J Med, 366(24), 2294–2304 (2012).
5) Psachoulia K, Chamberlain KA, Heo D, Davis SE, Paskus JD, Nanescu SE, Dupree JL, Wynn TA, Huang JK. IL4I1 augments CNS remyelination and axonal protection by modulating T cell driven inflammation. Brain, 139(Pt 12), 3121–3136 (2016).
6) Yamazaki R, Baba H, Yamaguchi Y. Unconventional Myosin ID is Involved in Remyelination After Cuprizone-Induced Demyelination. Neurochem Res, 43(1), 195–204 (2018).
7) Blakemore WF, Franklin RJ. Remyelination in experimental models of toxin-induced demyelination. Curr Top Microbiol Immunol, 318, 193–212 (2008).
8) Keough MB, Jensen SK, Yong VW. Experimental demyelination and remyelination of murine spinal cord by focal injection of lysolecithin. J Vis Exp, (97), 52679 (2015).
9) Weeks BA, Keisler AS, Myrvik QN, Warinner JE. Differential uptake of neutral red by macrophages from three species of estuarine fish. Dev Comp Immunol, 11(1), 117–124 (1987).
10) Ohkuma S, Poole B. Fluorescence probe measurement of the intralysosomal pH in living cells and the perturbation of pH by various agents. Proc Natl Acad Sci USA, 75(7), 3327–3331 (1978).
11) Baydyuk M, Cha DS, Hu J, Yamazaki R, Miller EM, Smith NV, Kelly KA, Huang JK. Tracking the evolution of CNS remyelinating lesion in mice with neutral red dye. Proc Natl Acad Sci USA, 116(28), 14290–14299 (2019).
12) Yamazaki R, Osanai Y, Kouki T, Shinohara Y, Huang JK, Ohno N. Macroscopic detection of demyelinated lesions in mouse PNS with neutral red dye. Sci Rep, 11(1), 16906 (2021).
13) Blasi F, Whalen MJ, Ayata C. Lasting pure-motor deficits after focal posterior internal capsule white-matter infarcts in rats. J Cereb Blood Flow Metab, 35(6), 977–984 (2015).
14) Lee MA, Blamire AM, Pendlebury S, Ho KH, Mills KR, Styles P, Palace J, Matthewa PM. Axonal injury or loss in the internal capsule and motor impairment in multiple sclerosis. Arch Neurol, 57(1), 65–70 (2000).
15) Maimone D, Reder AT, Finocchiaro F, Recupero E. Internal capsule plaque and tonic spasms in multiple sclerosis. Arch Neurol, 48(4), 427–429 (1991).
16) Schiemanck SK, Kwakkel G, Post MW, Kappelle LJ, Prevo AJ. Impact of internal capsule lesions on outcome of motor hand function at one year post-stroke. J Rehabil Med, 40(2), 96–101 (2008).
17) Shelton FN, Reding MJ. Effect of lesion location on upper limb motor recovery after stroke. Stroke, 32(1), 107–112 (2001).
18) Yamazaki R, Ohno N, Huang JK. Acute motor deficit and subsequent remyelination-associated recovery following internal capsule demyelination in mice. J Neurochem, 156(6), 917–928 (2021).

This page was created on 2022-11-04T14:39:16.136+09:00
This page was last modified on 2023-01-11T13:55:46.000+09:00
このサイトは(株)国際文献社によって運用されています。