脳内マクロファージの統合的理解に向けた研究
九州大学 生体防御医学研究所 分子神経免疫学分野
発行日:2023年12月30日Published: December 30, 2023
© 2023 日本神経化学会© 2023 The Japanese Society for Neurochemistry
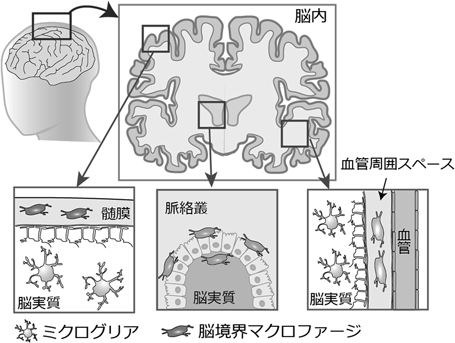
脳や脊髄といった中枢神経系が果たす高度な機能は、神経細胞やグリア細胞、血管系細胞など多様な細胞によって維持されている。中枢神経系の主要免疫細胞として知られるミクログリアは、1919年にスペイン人神経科学者のPio del Rio-Hortega博士によって記述されて以降、様々な方法でその機能解析が進められてきた(図1)。それらは、死細胞の除去や組織炎症の制御といったいわゆる免疫細胞としての機能に加え、神経新生・保護や髄鞘形成の促進といった脳形成過程への関与など、非免疫機能を含めた多様な脳内生理現象に関わると考えられている1)。一方、脳髄膜や血管周囲スペース(Virchow-Robinスペースとも呼ぶ)、脳脊髄液の産生を担う脈絡叢といった中枢と末梢の境界領域には、脳境界マクロファージ(CAM: CNS border-associated macrophage)と呼ばれるミクログリアとはタイプの異なるマクロファージが存在する(図1)2)。本稿では、1細胞解析やFate mappingなどの最新技術を用いて、我々が進めてきた脳内マクロファージに関する研究成果について概説する。
ミクログリアの機能や細胞特性が次々に明らかになっていくなかで、長らく議論が続いてきたのがミクログリアの多様性である。歴史的に、ミクログリアは、細胞形態や分布密度、電気生理学的特性や免疫関連分子の細胞膜上発現分子パターンなど、さまざま指標によって分類されてきた3)。そのような中、最近技術革新を遂げた1細胞解析技術は、ミクログリアの多様性に関する議論にブレークスルーを齎した。我々は、発達各時期のマウスの各脳領域からミクログリアを回収し、1細胞RNAシークエンシング(scRNA-seq)法を用いて多次元遺伝子発現解析を行った4)。その結果、胎生期の脳内には多様な遺伝子プロファイルを有したミクログリアが存在し、そうした多様性は脳の発達とともに減少していくことが明らかになった4)。また、成体脳内のミクログリアは比較的均一な細胞集団として存在し、脳領域特異的なミクログリア亜集団は存在しないことも分かった。一方、顔面神経切断モデルおよびCuprizone投与脱髄モデルという異なる疾患モデルマウスの病巣部から回収したミクログリアを同時比較解析することにより、各疾患モデル内で変化を遂げたミクログリアは、それぞれ全く異なる遺伝子発現パターンを有していることが明らかになった4)。特に、脱髄モデル由来のミクログリアは、病態の進行に伴って遺伝子発現パターンが変遷することも示され、ミクログリアが常に組織環境の変化に伴って、柔軟に遺伝子発現パターンの変化させていることが示された。一方、病態モデル内で機能的変化を遂げたミクログリアは、その後どういった運命を辿るのか?顔面神経損傷モデルマウスにおいて、損傷神経の細胞体が存在する顔面神経核付近ではミクログリアが増殖を伴って集積し、同時に遺伝子発現パターンも劇的に変化する4, 5)。興味深いことに、神経損傷後しばらくすると、集積していたミクログリアの一部は細胞死を起こし、また残りの細胞は病巣部周囲の領域へと拡散していき、正常時の細胞分布密度へと回帰することが明らかになった。またその際には、遺伝子発現パターンも元の状態に戻っていく。つまり、ミクログリアは高度な可塑性を持ち、そうした細胞特性を利用して華麗に細胞機能を変化させていることが明らかになった。
マウス等のモデル動物を用いた解析に加え、近年ヒトミクログリアを用いた各種オミクス解析が世界中で進められており、マウスミクログリアとの類似性や独自性が明らかになってきている6)。我々は、病理所見の見られない“healthy”な脳領域および難治性脱髄疾患として知られる多発性硬化症の患者脳内から回収したヒトミクログリアのscRNA-seq解析を世界で初めて行った4)。その結果、病理所見の見られない脳領域においては、比較的均一な細胞集団として存在するミクログリアが捉えられた一方で、多発性硬化症患者脳内では、アポリポプロテインEや転写因子MAFBを発現増加したミクログリアが観察され、それらはカテプシンDやアポリポプロテインC1を高発現するタイプ、MHCクラスII関連遺伝子(CD74など)を高発現するタイプ、さらにはオステオポンチンやリポプロテインリパーゼを高発現するタイプへと分類された4)。一方、脳腫瘍患者の脳・腫瘍組織内におけるミクログリアも高度な多様性を示しており、特に血管内皮細胞増殖因子Aを高発現するタイプやCD163を高発現するタイプは、脳腫瘍という組織環境が作り出す特殊なタイプであると考えられる7)。今後、病態ごとに異なるミクログリア亜集団を正確に捉え、その機能や分布パターン等を理解していく必要がある。
CAMsは、ミクログリアと同様に、胎生早期にyolk sac (YS)のblood island内に出現するerythro-myeloid progenitors (EMPs)を起源としていると考えられている8)。EMPsは、YS内で未成熟のA1細胞、そして前成熟A2細胞というマクロファージ前駆細胞へと分化した後、胎児の循環系を介して脳周囲に到達し、実質内へと浸潤しミクログリアへ、そして髄膜において髄膜マクロファージへと分化成熟する9)。しかし、ミクログリアとCAMsは、どの段階でそれぞれの細胞種に分かれるのか不明であった。我々は、YS前駆細胞の段階で細胞の運命決定が起きているのではないか、つまり、ミクログリアおよびCAMs特異的なYS前駆細胞が存在するのではないかと考えた。そこで、YSからA1・A2マクロファージ前駆細胞を回収し、scRNA-seq解析技術を用いて網羅的遺伝子発現解析を行った10)。その結果、yolk sac内に遺伝子発現プロファイルの異なる多様なマクロファージ前駆細胞が存在することが明らかになった。特に、PtprcやCx3cr1を高発現する、より成熟したA2前駆細胞は単一の細胞集団ではなく、Mrc1陽性の集団と陰性の集団として存在することが明らかになった10)。これまで、Mrc1はCAMsのマーカー分子として知られる因子であることから、ミクログリアとCAMsはYS内で既にその系統が分かれているという可能性、つまりMrc1陰性前駆細胞がミクログリアへ、Mrc1陽性前駆細胞がCAMsへと脳内で分化成熟する可能性が考えられた。この仮説の真偽を確かめるべく、タモキシフェン投与によってMrc1陽性前駆細胞を恒常的に蛍光タンパク質tdTomato標識できるMrc1CreERT2Rosa26tdTomatoマウスを用いて系譜トレーシング解析を行った。その結果、出生のMrc1CreERT2Rosa26tdTomatoマウス脳内には、tdTomato陽性のミクログリアおよびCAMsが観察され、予想に反して、Mrc1陽性前駆細胞はミクログリアにもCAMsにも分化する共通の前駆細胞亜集団であることが明らかになった10)。つまり、YS内にはマクロファージ前駆細胞の多様性が存在するものの、それらは必ずしもその後の細胞系譜を決定づけられた特異的な前駆細胞ではないということを意味している。
一方、胎生期の脳に定着した後のCAMsの挙動はほとんど明らかになっていなかった。そこで我々は、免疫組織染色やFate mapping技術等を用いて、髄膜および血管周囲マクロファージの分布動態を詳細に解析することにした10)。血管周囲マクロファージとは、血管内皮細胞やMural細胞等によって形成される基底膜と、アストロサイトの足底部に形成される基底膜によって挟まれた領域(血管周囲スペース)に存在するCAMsとして定義される。これまで、胎生期の脳内にすでに血管周囲マクロファージが存在すると考えられてきた。しかし、詳細な解析の結果、実は胎生期の脳皮質領域には血管周囲スペースは存在せず、出生前後に初めて狭小の血管周囲スペースが形成される10)。さらに、数日かけて十分な広さを持った血管周囲スペースへと成熟した後には、なんと髄膜に存在したマクロファージ(もしくは前駆細胞)が血管周囲スペースに遊走・定着し、血管周囲マクロファージになるが明らかになった。つまり、発達期の脳髄膜は、血管周囲マクロファージ前駆細胞の維持を担うニッチとして重要な役割を果たすことが明らかになった。
次に、髄膜からの細胞遊走および血管周囲へのマクロファージの定着に関わる脳境界構成細胞の特定を試みた。その結果、血管平滑筋細胞に異常をきたす遺伝子改変マウス(Notch3欠損)の脳内では、血管周囲マクロファージの数が極端に少なくなることが分かった10)。どのようなシグナル分子が、血管平滑筋細胞と血管周囲マクロファージ間の相互作用を仲介するのかは未だ明らかになっていない。また、血管周囲マクロファージの多くが、動脈周囲の血管周囲スペースに局在することも明らかになった10)。今後、その血管機能制御における役割について詳細に解析していく必要がある。
近年、神経免疫領域は非常に勢いがあり、脳内マクロファージに関する理解は、ここ数年で急速に進んでいる。本研究によって、長年議論が続いてきたミクログリアの多様性および可塑性に関する新たな概念を提唱するに至った。また、研究が先行しているミクログリアに続く脳内免疫システムにおける新たなプレイヤーとして、脳境界マクロファージに関してその発生、分布動態、遺伝子発現プロファイルといった基盤データの構築に至った。その機能や細胞特性がほとんど分かっていない今、どのように脳内免疫系に関与するのか想像の域を超えないが、脳境界領域において他の免疫細胞との相互作用を介して、様々な重要機能を担っていることが想定される。今後、ミクログリアと脳境界マクロファージの解析を同時に進めることで、脳の形成維持や疾患発症メカニズムに関する新たな概念の創出を目指したい。
本研究成果の多くは、フライブルク大学Marco Prinz教授の指揮のもと実施されました。Prinz教授をはじめ本研究に協力していただきました多くの共同研究者に深く感謝いたします。また、2023年度日本神経化学会優秀賞に選んでいただき、心より感謝申し上げます。
1) Prinz M, Jung S, Priller J. Microglia Biology: One Century of Evolving Concepts. Cell, 179(2), 292–311 (2019).
2) Masuda T. Recent topics regarding macrophage in the central nervous system. J Biochem, (2022).
3) Masuda T, Sankowski R, Staszewski O, Prinz M. Microglia Heterogeneity in the Single-Cell Era. Cell Rep, 30(5), 1271–1281 (2020).
4) Masuda T, Sankowski R, Staszewski O, Böttcher C, Amann L, Sagar, Scheiwe C, Nessler S, Kunz P, van Loo G, Coenen VA, Reinacher PC, Michel A, Sure U, Gold R, Grün D, Priller J, Stadelmann C, Prinz M. Spatial and temporal heterogeneity of mouse and human microglia at single-cell resolution. Nature, 566(7744), 388–392 (2019).
5) Tay TL, Mai D, Dautzenberg J, Fernández-Klett F, Lin G, Sagar, Datta M, Drougard A, Stempfl T, Ardura-Fabregat A, Staszewski O, Margineanu A, Sporbert A, Steinmetz LM, Pospisilik JA, Jung S, Priller J, Grün D, Ronneberger O, Prinz M. A new fate mapping system reveals context-dependent random or clonal expansion of microglia. Nat Neurosci, 20(6), 793–803 (2017).
6) Priller J, Prinz M. Targeting microglia in brain disorders. Science, 365(6448), 32–33 (2019).
7) Sankowski R, Bottcher C, Masuda T, Geirsdottir L, Sagar, Sindram E, Seredenina T, Muhs A, Scheiwe C, Shah MJ, Heiland DH, Schnell O, Grün D, Priller J, Prinz M. Mapping microglia states in the human brain through the integration of high-dimensional techniques. Nat Neurosci, 22(12), 2098–2110 (2019).
8) Kierdorf K, Masuda T, Jordao MJC, Prinz M. Macrophages at CNS interfaces: ontogeny and function in health and disease. Nat Rev Neurosci, 20(9), 547–562 (2019).
9) Kierdorf K, Erny D, Goldmann T, Sander V, Schulz C, Perdiguero EG, Wieghofer P, Heinrich A, Riemke P, Hölscher C, Müller DN, Luckow B, Brocker T, Debowski K, Fritz G, Opdenakker G, Diefenbach A, Biber K, Heikenwalder M, Geissmann F, Rosenbauer F, Prinz M. Microglia emerge from erythromyeloid precursors via Pu.1- and Irf8-dependent pathways. Nat Neurosci, 16(3), 273–280 (2013).
10) Masuda T, Amann L, Monaco G, Sankowski R, Staszewski O, Krueger M, Del Gaudio F, He L, Paterson N, Nent E, Fernández-Klett F, Yamasaki A, Frosch M, Fliegauf M, Bosch LFP, Ulupinar H, Hagemeyer N, Schreiner D, Dorrier C, Tsuda M, Grothe C, Joutel A, Daneman R, Betsholtz C, Lendahl U, Knobeloch KP, Lämmermann T, Priller J, Kierdorf K, Prinz M. Specification of CNS macrophage subsets occurs postnatally in defined niches. Nature, 604(7907), 740–748 (2022).

This page was created on 2023-11-06T15:34:20.592+09:00
This page was last modified on 2024-01-05T08:48:09.000+09:00
このサイトは(株)国際文献社によって運用されています。