思春期の社会経験依存的な眼窩前頭皮質-扁桃体回路のシナプス機能の変化
福井大学 子どものこころの発達研究センター 脳機能発達研究部門
発行日:2023年12月30日Published: December 30, 2023
© 2023 日本神経化学会© 2023 The Japanese Society for Neurochemistry
生後初期から発達期にかけての幼少期の時期は神経回路の可塑性が高く、経験依存的・神経活動依存的な機能的発達や再編成が盛んである1)。そのため、この時期の個体を取り巻く環境は、脳機能の発達に大きな影響を与えると考えられている。特に、幼少期の他個体との社会的相互作用は情動や認知機能の発達に重要であることが推測されており、サルやげっ歯類などの動物において、乳児期の母子間の相互作用あるいは思春期の他個体との社会的相互作用を剥奪すると、様々な認知機能や情動行動、社会性の異常がもたらされることが報告されている2–8)。また、ヒトにおいても、ネグレクトなどの経験はうつ病や一部のパーソナリティ障害などの精神疾患のリスクファクターとなることや、社会隔離環境下で育った児童は認知機能障害の罹患率が高いことが報告されている9, 10)。そのため、幼少期の社会的相互作用の不足が神経回路の機能やその発達に対して与える影響を紐解くことは、幼少期の環境が原因と推測される精神疾患症状の病態理解やその解決策を考えるうえで重要である。
筆者らは情動、社会性の制御に重要かつ幼少期の環境の影響を受ける脳領域として、前頭前皮質の腹側部領域に広がる眼窩前頭皮質(Orbitofrontal cortex: OFC)に注目し、研究を進めてきた。OFCは様々な感覚入力を受けることに加え、扁桃体や側坐核などの情動に密接に関連する領域と相互の神経連絡を持つ11)。OFCは情動処理や価値の評価、行動の柔軟性に関与しており12)、OFCを損傷した患者では様々な情動処理や社会性の異常が観察されることが報告されている13, 14)。加えて、幼少期のネグレクトなどが原因となりうる精神疾患で、OFCの活動の異常が観察されている15)。そのため、幼少期の社会的相互作用の剥奪が、OFCを中心とする神経回路の機能異常を引き起こし、様々な精神疾患症状の原因の一因となる可能性が考えられる。この可能性を明らかにするため、思春期マウスに対する隔離飼育による社会経験の剥奪が、OFCから扁桃体に投射するシナプスの機能に対して与える影響を検討した、筆者らの研究16)を本稿で紹介したい。
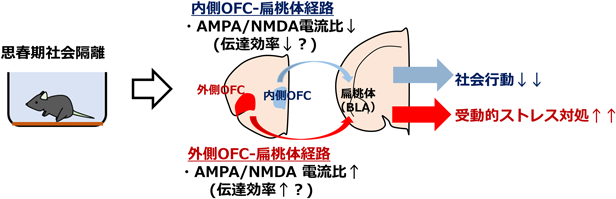
筆者らは、離乳後すぐの思春期のマウス(生後3週齢)に対し個別飼育を行うことで他個体との社会的相互作用を剥奪した(思春期社会隔離)。これらのマウスに対し、成熟後(生後8週齢~)に行動解析を行った結果、先行研究と同じく、他個体に対する社会行動の低下および強制水泳・尾懸垂試験における無動の増加などの受動的なストレス対処行動の増加が観察された。そこで、思春期社会隔離マウスにおいて、OFCから扁桃体外側基底核(BLA)に投射するシナプス機能の異常が観察されるか、スライスパッチクランプ法によるシナプス伝達記録により検討を行った。
OFCからBLAへの投射は複雑かつ長距離であり、電極を用いた局所刺激によってはシナプス伝達の単離記録が困難である。加えて、OFCは内側部・外側部などさらなる小領域に分かれており、小領域ごとの機能の違いが近年注目されている17, 18)。それぞれの小領域からBLAへの投射経路に対し、思春期社会隔離が異なる影響を与える可能性がある。そのため、光遺伝学とパッチクランプ記録を併用することで、内側・外側それぞれのOFC小領域から扁桃体BLA核へのシナプス伝達を単離計測した。まず、アデノ随伴ウィルス(AAV)ベクターを用いてマウスの内側OFCまたは外側OFCに限局して光感受性チャネルChR2を発現させた。その動物から投射先の扁桃体基底外側核(BLA)の急性スライスを作製し、錐体細胞からパッチクランプ記録下にて青色光を照射することで、それぞれのOFC小領域からBLAへ投射するシナプス伝達を単離計測できる。この系を用いて内側・外側OFC-BLA投射の興奮性シナプス伝達特性に対し、社会隔離が与える影響の評価を行った。
まず、プレシナプスの短期可塑性の指標であるpaired-pulse比について評価した結果、内側・外側いずれのOFC小領域からの投射シナプスにおいて、隔離による有意な変化は観察されなかった。次に、ポストシナプスの可塑性の指標の1つであるAMPA/NMDA電流成分比を評価した結果、隔離マウスでは内側OFC-BLAシナプスにおけるAMPA受容体由来の電流比の低下が観察された。一方で、外側OFC-BLAシナプスにおいては、隔離によってAMPA受容体電流比が増加するという反対の結果が観察された。これらの結果から、思春期の社会経験の剥奪はOFCから扁桃体への投射シナプスに対し、小領域ごとに異なるポストシナプスの機能的変化を引き起こすことが示された。AMPA受容体は興奮性伝達の早い成分の大部分を担うため、思春期の社会隔離により内側OFC-BLAでは興奮性シナプス伝達効率の低下、外側OFC-BLAシナプスでは増加が引き起こされている可能性が示唆される。
さらに筆者らは、光遺伝学的に行動下のマウスの外側・内側OFC-BLAシナプス伝達を操作し、社会隔離マウスで異常が見られた、社会行動・ストレス対処の制御に対するこれら経路のシナプス伝達の寄与を検討した。AAVベクターを用いてマウスの内側OFCまたは外側OFCにChR2あるいは抑制性陰イオンポンプNpHRを発現させ、投射先のBLAに埋め込んだLEDを介して光照射を行うことで、内側または外側OFCからBLAに投射するシナプス伝達の操作を行った。
まず、通常の集団飼育環境で生育したマウスの内側OFC-BLA投射のシナプス伝達を抑制した結果、社会隔離マウス同様に社会性が低下することが観察された。反対に、社会隔離マウスにおいてこの経路のシナプス伝達を活性化すると、隔離による社会性の低下の改善が観察された。次に、通常の集団飼育環境で生育したマウスにおいて外側OFC-BLAのシナプス伝達を活性化した結果、隔離マウス同様に尾懸垂試験の無動が増加することが観察された。反対に、隔離マウスにおいてこの経路のシナプス伝達を抑制すると、隔離による無動の増加が改善した。興味深いことに、内側OFC-BLAのシナプス伝達操作は無動、外側OFC-BLAの操作は社会行動にそれぞれ影響を与えなかった。これらの結果から、内側OFC-BLA経路のシナプス伝達は社会行動を、外側OFC-BLA経路のシナプス伝達は受動的ストレス対処行動をそれぞれ独立的に制御していることが示された。先述の電気生理学的解析の結果と合わせて考えると、思春期の社会隔離は内側・外側OFC-BLA経路に対して異なる機能的変化を引き起こすことで、別々の行動特性の変化に寄与することが示唆される(図1)。
筆者らの研究結果は内側・外側OFCから扁桃体に投射する神経回路が、幼少期の社会的な相互作用の不足が原因となる精神疾患症状の治療標的となる可能性を示唆している。近年、内側OFCまたは外側OFCを標的とした経頭蓋直流刺激や経頭蓋磁気刺激が、強迫性障害や抑うつ症状の改善に有用な例が報告されており19, 20)、これらの介入法の幼少期の環境が原因となる疾患への応用が期待される。
また、扁桃体は恐怖などのネガティブな感情価だけでなく、報酬行動や社会性といった多様な行動の制御や情報処理に対しても関与することが明らかにされている21–23)。内側・外側OFCは扁桃体内の同一の核(BLA)に投射しているが、それぞれの領域からの入力がBLA内でどのように情報処理を行い、社会性や受動的ストレス対処といった別々の行動を独立的に制御するのか、その回路メカニズムは興味深い。加えて、思春期の社会隔離がどのように内側・外側OFCからBLAへの投射に対し異なる機能的変化を引き起こすのか、その可塑性メカニズムも不明であるため、今後さらなる研究の展開によって解明していきたい。
本研究成果は、国立精神・神経医療研究センターで得られたものです。多大なるご指導を賜りました山田光彦先生(現東京家政学院大学)、関口正幸先生(現東京理科大学)、和田圭司先生に厚く御礼申し上げます。ならびに、現職の福井大学でご指導いただいている松﨑秀夫先生と学部・大学院生時代に、研究者として一からご指導いただいた鳥取大学生命科学科の一坂吏志先生、畠義郎先生に心から感謝申し上げます。また、私は修士・博士課程在籍時から、神経化学会の若手研究者育成セミナーにたびたび参加させていただきました。それに加え、本稿の執筆機会を与えて下さいました日本神経化学会および、優秀賞・奨励賞選考委員、編集委員委員の先生方に深く感謝申し上げます。今後も日本神経化学会奨励賞受賞者としての自覚を持ち、より一層精進して研究に取り組みたいと考えております。今後ともご指導ご鞭撻のほど、よろしくお願い申し上げます。
1) Hensch TK. Critical period plasticity in local cortical circuits. Nat Rev Neurosci, 6(11), 877–888 (2005).
2) Harlow HF, Dodsworth RO, Harlow MK. Total social isolation in monkeys. Proc Natl Acad Sci USA, 54(1), 90–97 (1965).
3) Francis DD, Diorio J, Plotsky PM, Meaney MJ. Environmental enrichment reverses the effects of maternal separation on stress reactivity. J Neurosci, 22(18), 7840–7843 (2002).
4) Kuniishi H, Ichisaka S, Yamamoto M, Ikubo N, Matsuda S, Futora E, Harada R, Ishihara K, Hata Y. Early deprivation increases high-leaning behavior, a novel anxiety-like behavior, in the open field test in rats. Neurosci Res, 123, 27–35 (2017).
5) Mogi K, Ishida Y, Nagasawa M, Kikusui T. Early weaning impairs fear extinction and decreases brain-derived neurotrophic factor expression in the prefrontal cortex of adult male C57BL/6 mice. Dev Psychobiol, 58(8), 1034–1042 (2016).
6) Makinodan M, Rosen KM, Ito S, Corfas G. A critical period for social experience-dependent oligodendrocyte maturation and myelination. Science, 337(6100), 1357–1360 (2012).
7) Tulogdi A, Tóth M, Barsvári B, Biró L, Mikics E, Haller J. Effects of resocialization on post-weaning social isolation-induced abnormal aggression and social deficits in rats. Dev Psychobiol, 56(1), 49–57 (2014).
8) Sargin D, Oliver DK, Lambe EK. Chronic social isolation reduces 5-HT neuronal activity via upregulated SK3 calcium-activated potassium channels. eLife, 5, e21416 (2016).
9) Behen ME, Helder E, Rothermel R, Solomon K, Chugani HT. Incidence of specific absolute neurocognitive impairment in globally intact children with histories of early severe deprivation. Child Neuropsychol, 14(5), 453–469 (2008).
10) Naughton AM, Maguire SA, Mann MK, Lumb RC, Tempest V, Gracias S, Kemp AM. Emotional, behavioral, and developmental features indicative of neglect or emotional abuse in preschool children: a systematic review. JAMA Pediatr, 167(8), 769–775 (2013).
11) Rolls ET. The functions of the orbitofrontal cortex. Brain Cogn, 55(1), 11–29 (2004).
12) Rempel-Clower NL. Role of orbitofrontal cortex connections in emotion. Ann N Y Acad Sci, 1121(1), 72–86 (2007).
13) Beer JS, John OP, Scabini D, Knight RT. Orbitofrontal cortex and social behavior: integrating self-monitoring and emotion-cognition interactions. J Cogn Neurosci, 18(6), 871–879 (2006).
14) Hornak J, Bramham J, Rolls ET, Morris RG, O’Doherty J, Bullock PR, Polkey CE. Changes in emotion after circumscribed surgical lesions of the orbitofrontal and cingulate cortices. Brain, 126(Pt 7), 1691–1712 (2003).
15) Hornak J, Bramham J, Rolls ET, Morris RG, O’Doherty J, Bullock PR, Polkey CE. Changes in emotion after circumscribed surgical lesions of the orbitofrontal and cingulate cortices. Brain, 126(Pt 7), 1691–1712 (2003).
16) Kuniishi H, Nakatake Y, Sekiguchi M, Yamada M. Adolescent social isolation induces distinct changes in the medial and lateral OFC-BLA synapse and social and emotional alterations in adult mice. Neuropsychopharmacology, 47(9), 1597–1607 (2022).
17) Cheng W, Rolls ET, Qiu J, Liu W, Tang Y, Huang CC, Wang X, Zhang J, Lin W, Zheng L, Pu J, Tsai SJ, Yang AC, Lin CP, Wang F, Xie P, Feng J. Medial reward and lateral non-reward orbitofrontal cortex circuits change in opposite directions in depression. Brain, 139(Pt 12), 3296–3309 (2016).
18) Izquierdo A. Functional Heterogeneity within Rat Orbitofrontal Cortex in Reward Learning and Decision Making. J Neurosci, 37(44), 10529–10540 (2017).
19) Grover S, Nguyen JA, Viswanathan V, Reinhart RMG. High-frequency neuromodulation improves obsessive-compulsive behavior. Nat Med, 27(2), 232–238 (2021).
20) Feffer K, Fettes P, Giacobbe P, Daskalakis ZJ, Blumberger DM, Downar J. 1 Hz rTMS of the right orbitofrontal cortex for major depression: Safety, tolerability and clinical outcomes. Eur Neuropsychopharmacol, 28(1), 109–117 (2018).
21) Namburi P, Beyeler A, Yorozu S, Calhoon GG, Halbert SA, Wichmann R, Holden SS, Mertens KL, Anahtar M, Felix-Ortiz AC, Wickersham IR, Gray JM, Tye KM. A circuit mechanism for differentiating positive and negative associations. Nature, 520(7549), 675–678 (2015).
22) Beyeler A, Chang CJ, Silvestre M, Lévêque C, Namburi P, Wildes CP, Tye KM. Organization of Valence-Encoding and Projection-Defined Neurons in the Basolateral Amygdala. Cell Rep, 22(4), 905–918 (2018).
23) Fustinana MS, Eichlisberger T, Bouwmeester T, Bitterman Y, Luthi A. State-dependent encoding of exploratory behaviour in the amygdala. Nature, 592(7853), 267–271 (2021).

This page was created on 2023-11-06T17:36:42.749+09:00
This page was last modified on 2024-01-05T08:50:53.000+09:00
このサイトは(株)国際文献社によって運用されています。