軸索再伸長を基盤としたアルツハイマー病の根本的治療戦略の開発
富山大学 和漢医薬学総合研究所 神経機能学領域
発行日:2023年12月30日Published: December 30, 2023
© 2023 日本神経化学会© 2023 The Japanese Society for Neurochemistry
近年、認知症患者数の急増は深刻な社会問題となっており、2025年には高齢者の5人に1人が認知症に罹患すると推測されている。認知症の過半数を占めるのが、アルツハイマー病(AD)である。ADでは、脳にamyloid β (Aβ)が沈着することで、様々な神経機能を担う神経回路網が破綻し、認知機能障害を始めとした脳機能障害が引き起こされる。しかし、長年承認薬として用いられてきたコリンエステラーゼ阻害薬(ドネペジル、ガランタミン、リバスチグミン)やNMDA受容体拮抗薬(メマンチン)は、認知機能障害の進行を遅らせる対症療法にとどまっている。また、2023年に原因物質(Aβ)の除去を狙った新薬として、Aβ抗体のレカヌマブが各国で承認された。軽度認知障害または軽度ADの比較的早期のステージからレカヌマブを投与すると、脳内のAβが減少し認知機能障害の進行は数ヶ月遅れるものの、認知機能の低下が完全に止まるわけではないことが示されている1)。この理由は、AD発症よりも20–30年も前の無症状期からAβの蓄積が始まっており、それに伴って神経回路が進行性に変性するため、Aβの除去だけでは既に破綻した神経回路の修復は困難だからである。
そこで筆者は、“破綻した脳の神経回路を再形成する”ことが、ADの根本的治療に必須であると考えた。神経回路の形成には、神経細胞の軸索が投射先の神経細胞の樹状突起にシナプス結合する必要があるが、特に軸索は遠く離れた脳部位に向かっても長く伸長しなければならない。しかし、成体の脳においては、一度変性した軸索は再伸長できないと長年考えられてきたため、治療戦略としてほとんど着目されてこなかった。近年、成体脳においても軸索が近位には自発的に再伸長することが報告されたものの2, 3)、軸索が遠く離れた投射先に向かって再伸長するかどうかを評価するには至っておらず、これを達成できる薬物も見出されてこなかった。本稿では、ADモデルマウス脳内の軸索を長距離に向かって再伸長させ、記憶障害を回復させた和漢薬成分“diosgenin”の研究成果について紹介する。
筆者の研究グループでは、かねてより、初代培養神経細胞においてAβで萎縮した軸索を再伸長させる薬物を探索してきた。その中で、特に優れた軸索再伸長活性を示したのが、山薬(ヤマノイモ科Dioscorea japonicaまたはD. batatasの根茎)中に含有されるステロイドサポゲニン化合物のdiosgeninである(図1)。また、diosgeninをADモデル(5XFAD)マウスに投与し、記憶障害の回復作用を検討した。5XFADマウスとは、5種類の家族性アルツハイマー病変異遺伝子を神経細胞特異的に過剰発現させたトランスジェニックマウスである4)。このマウスでは、約2ヶ月齢から脳でAβの蓄積が始まり、4–5ヶ月齢から記憶障害が生じる。記憶障害が進行した6–8ヶ月齢の5XFADマウスにdiosgeninを20日間投与すると、マウスの物体認知記憶障害が回復した5)。
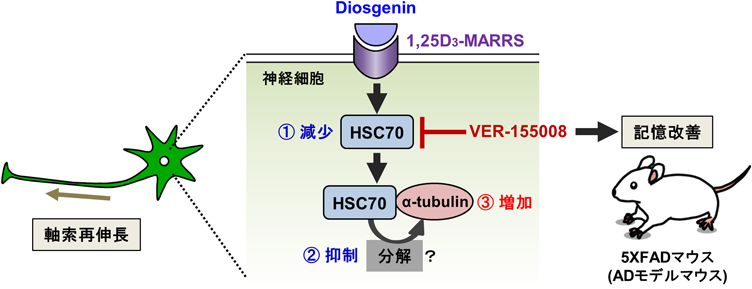
Diosgeninの軸索再伸長及び記憶回復に関わる直接結合タンパク質をdrug affinity responsive target stability (DARTS)法で網羅的に探索したところ、活性型ビタミンD3受容体である1,25D3-membrane-associated rapid response steroid-binding receptor (1,25D3-MARRS)が同定された5, 6)。また、diosgeninによる1,25D3-MARRSの刺激後に神経細胞内で引き起こされる分子シグナル変化を解析した結果、野生型マウスと比べて5XFADマウスの脳では増加するが、diosgeninを投与した5XFADマウスの脳で減少したタンパク質として、heat shock cognate (HSC) 70を同定した。HSC70の活性阻害剤であるVER-155008を初代培養神経細胞に処置すると、Aβで萎縮した軸索が再伸長した。また、VER-155008を5XFADマウスに投与すると、マウスの物体認知記憶、空間記憶、エピソード記憶のいずれの障害も改善され、脳内の軸索終末の変性やAβプラーク、リン酸化タウの蓄積も減少した。この際、投与されたVER-155008が脳に移行していることも確認された7)。さらに、HSC70の発現量が減少した後に、軸索再伸長に直接つながる下流分子を探索した結果、神経細胞中でHSC70と結合するタンパク質として、細胞骨格タンパク質であるα-tubulinを同定した。HSC70は、様々な結合タンパクの分解を促進するが、Aβを処置した軸索上ではα-tubulinが減少した。一方、Aβ処置後からdiosgeninを処置すると、軸索上のα-tubulin量が正常レベルにまで増加した。よって、diosgeninはHSC70を減少させることで、HSC70によるα-tubulinの分解を抑制し、軸索再伸長や記憶回復に関わる可能性が示された(図2)8)。
Diosgenin投与が5XFADマウス脳内の萎縮した軸索をつながるべき脳部位に再伸長させるかどうかを検討するために、記憶形成に関わる海馬–前頭前野に着目した9)。野生型マウス及び5XFADマウスの前頭前野に、赤色蛍光の逆行性トレーサーDextran Texas Red (3000 MW)を注入後、溶媒またはdiosgeninを14日間経口投与し、同じ前頭前野に緑色蛍光の逆行性トレーサーDextran FITC (3000 MW)を注入した。その7日後に脳を摘出し、海馬切片を観察した。薬物投与によって海馬–前頭前野において軸索が萎縮した神経細胞は赤色のみで、軸索が再伸長した神経細胞は緑色のみで、軸索が伸びも縮みもしなかった神経細胞は赤・緑色の2色で標識された。解析の結果、5XFADマウスでは海馬–前頭前野に投射する軸索が減少していること、また、diosgeninの投与により本回路の軸索が再伸長することがそれぞれ示された10)。続いて、本回路において軸索が再伸長した海馬神経細胞をレーザーマイクロダイセクションによって採取し、発現量が変化する遺伝子を網羅的に探索した。その結果、軸索が再伸長した神経細胞において発現量が最も増加したタンパク質として、SPARC (Secreted protein acidic rich in cysteine)及びGalectin-1 (lectin, galactose binding soluble 1)を同定した。
AAV9ベクターを用いて海馬神経細胞特異的にSPARCまたはGalectin-1を過剰発現すると、5XFADマウスの物体認知記憶及び空間記憶の障害が回復し、海馬から前頭前野に投射する軸索の再伸長も促進された。また、DREADD (designer receptors exclusively activated by designer drugs)システムを用いて、5XFADマウスの海馬から前頭前野に軸索を再投射した神経細胞特異的に、その興奮性活動を抑制すると、SPARCの過剰発現によっていったん示された記憶改善作用が消失した10)。さらに、軸索伸長中の軸索では特に軸索膜上でSPARCが増加すること、軸索の軌跡に沿うように細胞外I型collagenが並ぶこと、軸索が萎縮してもI型collagenは残ること、軸索膜上のSPARCは細胞外に残存したI型collagenをたどるように相互作用することで、元々伸びていた方向に軸索が再伸長することを明らかにした10)。また、Galectin-1は海馬神経細胞の成長円錐膜上で発現し、前頭前野から分泌される新規軸索誘導因子Secernin-1の受容体としてはたらくことで、海馬–前頭前野の軸索誘導に関わることを示した11)。以上より、diosgeninがAD脳内の軸索を長距離に再伸長させることを明らかにし、本現象を担う責任分子としてSPARC及びGalectin-1を同定した(図3)。
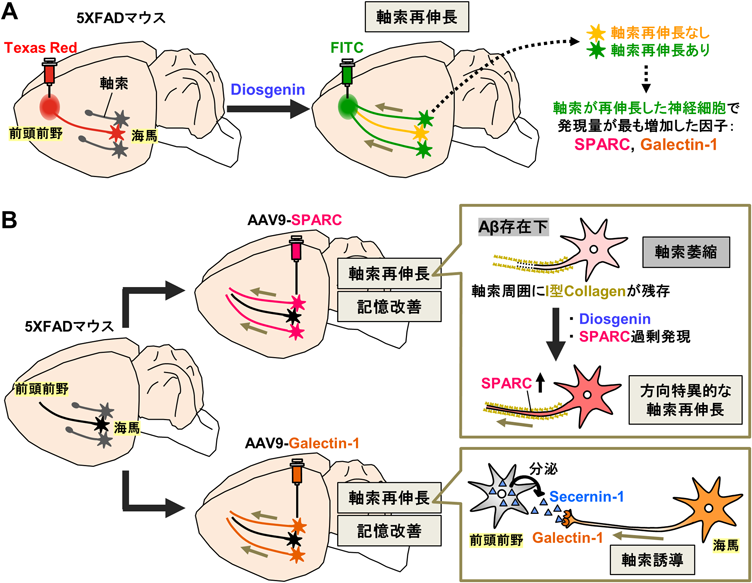
(A) 5XFADマウスにdiosgeninを投与すると、海馬–前頭前野において軸索が再伸長した。(B)海馬–前頭前野での軸索再伸長に関わる神経細胞中の機能分子として、SPARC及びGalectin-1を同定した。
ここまでの研究成果より、diosgeninがADに対して有力な新規治療薬候補であることを提示してきた。Diosgeninの基礎研究を社会実装するにあたり、臨床研究の実行可能性の観点から、化合物diosgeninでの創薬だけでなく、diosgeninを含有した山薬エキスでのトランスレーショナルリサーチを進めることにした。山薬が食薬区分上の非医であることを生かし、これを機能性表示食品として開発することを視野に入れ、正常マウスに対する山薬エキスの記憶亢進作用を検討した。山薬中のdiosgenin含量は、Dioscorea属の種類に依存して大きく異なる。特に、日本薬局方で規定される山薬(基原植物がD. japonica又はD. batatas)は、diosgenin含量が非常に低い12)。一方で、山薬中には、種々のdiosgenin配糖体が含有されており、服用後に生体内(主に腸内細菌による)でdiosgeninに代謝されるとされているが13)、その代謝効率は低い14)。そこで、日本薬局方の山薬を常法通り熱水抽出した場合の常法山薬エキス、及び中国河南省で栽培されたdiosgenin配糖体の含量が高いD. batatasを基原とする山薬を含水エタノールで抽出後、酸加水分解処置を施し、エキス中のdiosgenin含量を15–16%まで高めたdiosgenin高濃度山薬エキスを用いて、正常マウスに対する記憶亢進作用を検討した。その結果、日本薬局方の山薬熱水抽出エキス中にはdiosgeninがほとんど含まれておらず、経口投与後に十分量のdiosgeninが脳移行しないため、記憶亢進作用を示さなかった。一方で、diosgenin高濃度山薬エキスを油溶媒で経口投与すると、diosgeninの脳移行が格段に向上し、マウスの記憶が亢進した15, 16)。
続いて、このdiosgenin高濃度山薬エキスを用いて、健常人に対する認知機能向上作用を検討することを目的として、臨床試験を行った。20–81歳の28名の被験者に対し、ランダム化二重盲検クロスオーバー試験にてプラセボまたは山薬エキスの12週間服用前後で、認知機能を測るRepeatable Battery for the Assessment of Neuropsychological Status(RBANS)試験を行った。その結果、プラセボ群と比較して山薬エキスを12週間服用した群では、認知機能が向上することが示された17)。現在は、軽度認知障害及び軽度AD患者の認知機能に対する臨床試験を実施し終え、結果解析中である。さらに、これらの科学的根拠に基づき、機能性表示食品「ジオスゲニン・ゴールド(レジリオ株式会社)」の開発に繋げることができた。
成体脳では、損傷した神経回路は二度と再生できないと長年認識されてきたため、ADを始めとした神経変性疾患の根本的治療は不可能であると一般的に考えられてきた。しかし、diosgeninの発見により、AD脳の破綻した神経回路を再形成し、脳機能を再び活性化しうることを示すことができた。病因物質に着目しADの発症や進行を遅らせるという治療戦略に加えて、発症後の機能障害の回復が実現してこそ、真の根本的治療となる。今後も、diosgeninをはじめ神経回路の再形成に有用な薬物の作用メカニズム解析を展開し、脳の神経回路が修復される分子機構のさらなる解明を目指していきたい。
本研究を行うにあたり、学部生時代から現在に至るまで、様々な面から親身なご指導を賜りました富山大学和漢医薬学総合研究所神経機能学領域 東田千尋教授に、この場をお借りして心より厚く御礼申し上げます。また、日々支えてくださいました研究室の皆様、多くの共同研究者の先生方にも心より感謝申し上げます。
今でも、学部生の時に初めて日本神経化学会大会の若手道場セッションで発表させていただいた緊張感を鮮明に覚えています。日本神経化学会の先生方には、多くの刺激と成長の機会を与えていただき、深く感謝申し上げます。また、この度、日本神経化学会奨励賞を賜り、本稿の執筆機会を与えて下さいました日本神経化学会優秀賞・奨励賞選考委員の先生方、ならびに関係者の先生方、編集部の皆様に深く感謝申し上げます。
1) van Dyck CH, Swanson CJ, Aisen P, Bateman RJ, Chen C, Gee M, Kanekiyo M, Li D, Reyderman L, Cohen S, Froelich L, Katayama S, Sabbagh M, Vellas B, Watson D, Dhadda S, Irizarry M, Kramer LD, Iwatsubo T. Lecanemab in Early Alzheimer’s Disease. N Engl J Med, 388(1), 9–21 (2023).
2) Jin Y, Dougherty SE, Wood K, Sun L, Cudmore RH, Abdalla A, Kannan G, Pletnikov M, Hashemi P, Linden DJ. Regrowth of serotonin axons in the adult mouse brain following injury. Neuron, 91(4), 748–762 (2016).
3) Li S, Overman JJ, Katsman D, Kozlov SV, Donnelly CJ, Twiss JL, Giger RJ, Coppola G, Geschwind DH, Carmichael ST. An age-related sprouting transcriptome provides molecular control of axonal sprouting after stroke. Nat Neurosci, 13(12), 1496–1504 (2010).
4) Oakley H, Cole SL, Logan S, Maus E, Shao P, Craft J, Guillozet-Bongaarts A, Ohno M, Disterhoft J, Van Eldik L, Berry R, Vassar R. Intraneuronal beta-amyloid aggregates, neurodegeneration, and neuron loss in transgenic mice with five familial Alzheimer’s disease mutations: potential factors in amyloid plaque formation. J Neurosci, 26(40), 10129–10140 (2006).
5) Tohda C, Urano T, Umezaki M, Nemere I, Kuboyama T. Diosgenin is an exogenous activator of 1,25D3-MARRS/Pdia3/ERp57 and improves Alzheimer’s disease pathologies in 5XFAD mice. Sci Rep, 2(1), 535 (2012).
6) Tohda C, Lee YA, Goto Y, Nemere I. Diosgenin-induced cognitive enhancement in normal mice is mediated by 1,25D3-MARRS. Sci Rep, 3(1), 3395 (2013).
7) Yang X, Tohda C. Heat shock cognate 70 inhibitor, VER-155008, reduces memory deficits and axonal degeneration in a mouse model of Alzheimer’s disease. Front Pharmacol, 9, 48 (2018).
8) Yang X, Tohda C. Diosgenin restores Aβ-induced axonal degeneration by reducing the expression of heat shock cognate 70 (HSC70). Sci Rep, 8(1), 11707 (2018).
9) Wang C, Furlong TM, Stratton PG, Lee CCY, Xu L, Merlin S, Nolan C, Arabzadeh E, Marek R, Sah P. Hippocampus-prefrontal coupling regulates recognition memory for novelty discrimination. J Neurosci, 41(46), 9617–9632 (2021).
10) Yang X, Tohda C. Diosgenin restores memory function via SPARC-driven axonal growth from the hippocampus to the PFC in Alzheimer’s disease model mice. Mol Psychiatry, 28(6), 2398–2411 (2023).
11) Yang X, Tohda C. Axonal regeneration mediated by a novel axonal guidance pair, Galectin-1 and Secernin-1. Mol Neurobiol, 60(3), 1250–1266 (2023).
12) Vendl O, Wawrosch C, Noe C, Molina C, Kahl G, Kopp B. Diosgenin contents and DNA fingerprint screening of various yam (Dioscorea sp.) genotypes. Z Naturforsch C J Biosci, 61(11–12), 847–855 (2006).
13) Yi T, Fan LL, Chen HL, Zhu GY, Suen HM, Tang YN, Zhu L, Chu C, Zhao ZZ, Chen HB. Comparative analysis of diosgenin in Dioscorea species and related medicinal plants by UPLC-DAD-MS. BMC Biochem, 15(1), 19 (2014).
14) Okawara M, Tokudome Y, Todo H, Sugibayashi K, Hashimoto F. Enhancement of diosgenin distribution in the skin by cyclodextrin complexation following oral administration. Biol Pharm Bull, 36(1), 36–40 (2013).
15) Yang X, Nomoto K, Tohda C. Diosgenin content is a novel criterion to assess memory enhancement effect of Yam extracts. J Nat Med, 75(1), 207–216 (2021).
16) Tohda C, Yang X, Nomoto K. Transported amount of diosgenin to the brain is differed by a solvent fat. JJFCS, 27(2), 102–105 (2020).
17) Tohda C, Yang X, Matsui M, Inada Y, Kadomoto E, Nakada S, Watari H, Shibahara N. Diosgenin-rich yam extract enhances cognitive function: a placebo-controlled, randomized, double-blind, crossover study of healthy adults. Nutrients, 9(10), 1160 (2017).

This page was created on 2023-11-06T16:59:00.922+09:00
This page was last modified on 2024-01-05T08:56:47.000+09:00
このサイトは(株)国際文献社によって運用されています。