大脳皮質発生を分子・細胞・メタ細胞群の時空間ダイナミクスとして理解する
東京慈恵会医科大学解剖学講座・慶應義塾大学医学部解剖学教室
発行日:2023年12月30日Published: December 30, 2023
© 2023 日本神経化学会© 2023 The Japanese Society for Neurochemistry
大脳皮質が多様で高度な機能を有す背景には、発生における最初のステップとして、巧妙にニューロン移動が制御され、多くのサブタイプの興奮性ニューロンと抑制性ニューロンが整然とした層構造をとることで、複雑かつ精緻な回路形成が可能になった点を指摘できる。大脳皮質興奮性ニューロンは、脳室に面した脳室帯で分裂する神経幹細胞に由来する。そのまま分裂しないニューロンとなるものもあれば、脳表面に向かって移動し脳室下帯で分裂し、ニューロンを生み出すものもある1–3)。いずれの場合においても、最終的には、脳表面の辺縁帯直下で、先に移動を終えた早生まれのニューロンを追い越すターミナルトランスロケーションによって、早生まれのニューロンほど深層に、遅生まれのニューロンほど浅層に配置される(inside-outパターン)4, 5)。抑制性ニューロンは基底核原基で産生後、大脳皮質を主に水平方向に移動し、サブタイプにもよるがやはり誕生日依存的に配置される6)。
以前から、ニューロン移動の粗大な異常によって層構造が逆転することが、滑脳症などの重度な小児神経疾患の病態に重要な役割を果たしていることが知られている。しかし、神経発達の異常が病態の背景に想定されている多くの神経発達障害で、所見は重度な小児神経疾患よりも軽微であり7)、統合失調症などの精神神経疾患に至っては神経病理学者の墓場といわれてきたように再現性ある有用な所見もほとんどなく8)、具体的にどのような脳発生の異常があるかは間接的な観察所見を総合的に積み重ねることで推察している段階である9)。ニューロン移動を含めた、正常神経発生の精緻な制御を解明することは、死後脳研究や動物モデル、オルガノイドにおける所見を正しく解釈し、最終的にこうした障害の病態解明を進めていく上で必須である。
筆者はニューロン移動の制御は、分子による制御として理解すべきメカニズムと、細胞レベルでの制御として理解すべきものとがあると仮説を立てて、研究を進めてきた。本稿では、浅学非才を顧みず、こうした筆者の研究を中心に紹介したい。
はじめに、適切なニューロン移動がどのように分子によって制御されているか調べた10)。マウス大脳皮質表層ニューロンは、脳室帯で産生された後、その直上(多極性細胞蓄積帯、MAZ)で多数の突起を活発に伸縮させながら1日程度滞留するような振る舞い(多極性移動)を示す3, 11, 12)。その後ニューロンは水平方向に軸索伸張しつつ13–15)、放射状グリア線維をつたってシャクトリムシのように表面に移動する(ロコモーション)3, 16)。多極性移動は特異な移動様式であり、脆弱なステップである17)にもかかわらず、分子メカニズムは断片的な報告18)があるに過ぎなかった。筆者らはまず、子宮内電気穿孔法(EP)で移動ニューロンをラベルしてライブイメージングや組織学的解析を行った。多極性細胞の突起はF-actinに富んでおり、その突起は軸索成長突起のラメリポディアやフィロポディアと形態的に類似していることがわかった。そこでactinのリモデリングに関与するLpdやEna/Vaspファミリータンパク質に注目した。Lpdは細胞膜にあるphosphatidylinositol(3,4)-bisphosphate[PI(3,4)P2]と結合しつつ、Ena/Vaspをリクルートし、これがactinフィラメントの細胞膜へのアセンブリを制御することが主にin vitroの実験19–21)やC. elegansでの実験22)で示唆されていた。免疫組織化学とin situ hybridizationにより、LpdやMena(Ena/Vaspの一員)がMAZの多極性細胞で発現していることを示した。LpdやEna/Vaspの機能阻害によって多極性細胞の一次突起数が減じた。免疫染色やFörster共鳴エネルギー移動(FRET)で、多極性移動の突起先端にPI(3,4)P2が局在していることを示した。PI(3,4,5)P3をPI(3,4)P2に変換するShip2をノックダウンや特異的阻害剤により機能阻害したところ、やはり多極性細胞の一次突起数が減じた。こうした観察から、多極性細胞の特徴的な振る舞いには、PI(3,4)P2やLpd、Ena/Vaspが関与していることが示唆された(図1A)。また、このメカニズムが多極性移動の意義に関して重要な示唆を与えた。すなわち、多極性移動は横走する多数の軸索束をくぐり抜ける移動様式であるが、このようにLpdをノックダウンし(多極性移動に異常を起こさせて)ロコモーションすべき時期に観察したところ、移動ニューロンは脳表面に向かって移動せず、脳表面に平行に移動し続けた23)。軸索束という障害物をくぐり抜けて放射状線維という高速道路に正しくのるために、多数の突起を活発に伸縮させ環境を感知する必要があると考えた。
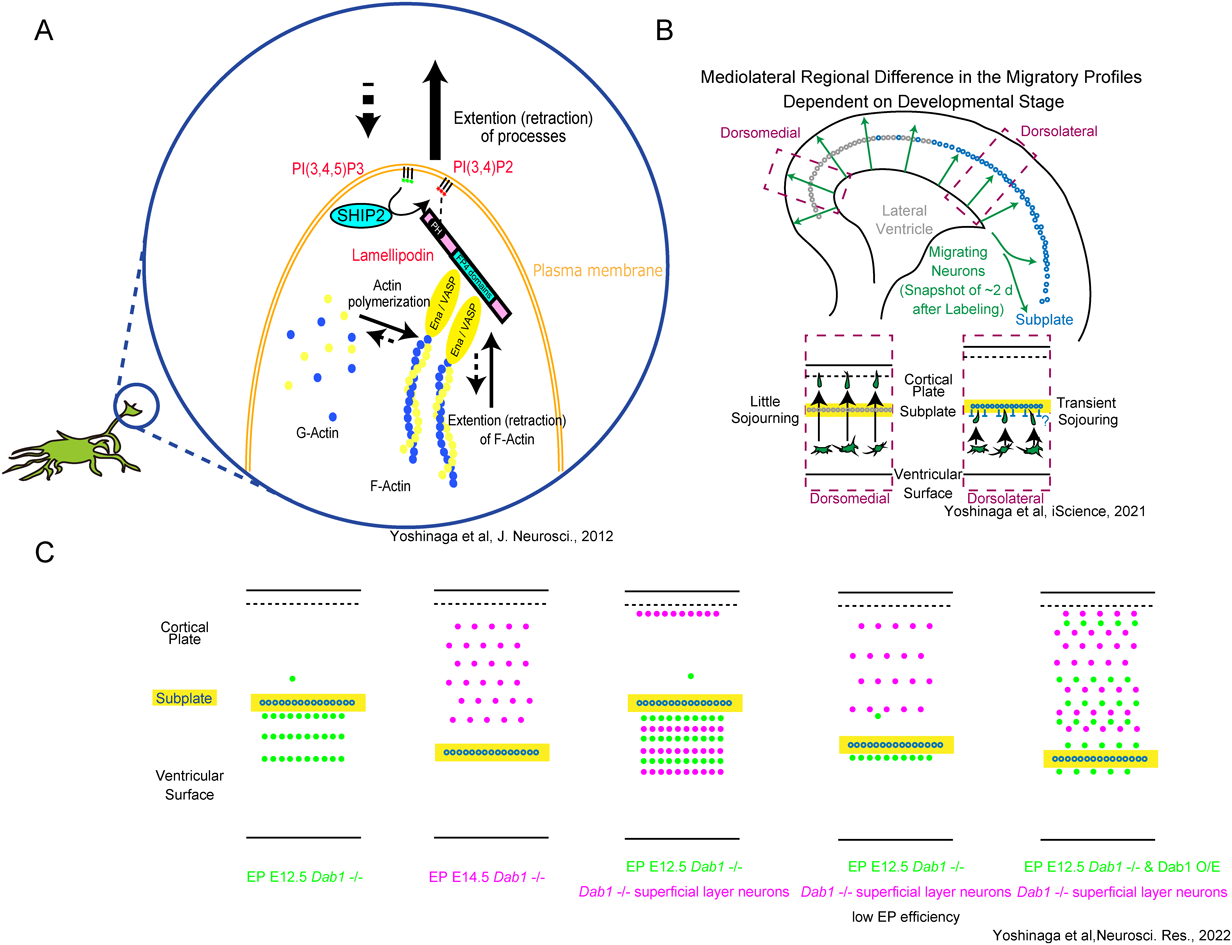
前述の研究から、移動ニューロンそのものの制御だけではなく、移動ニューロンと相互作用する細胞外環境も重要ではないかと考えた。マクロな発生の視点では、発生期大脳皮質の表面(辺縁帯)にはCajal-Retzius細胞がおり、Reelinを分泌しており、これを欠損する哺乳類個体では皮質層構造が逆転することが古くから知られている24–26)。しかし、筆者らは、さきの研究から、軸索やほかの細胞、放射状線維など、もっと構造が局所において相互作用するような移動制御に興味を持っていた。
まず、免疫組織化学により、最初期に産生・配置されるニューロン(サブプレートニューロン)や視床皮質投射線維には、領域により質的・量的な違いがあることに違いがあることに気づいた。しかしチミジンアナログを利用した研究から、皮質ニューロンの移動は、基底核を回り込む腹側を除いて、移動の領域差はほとんどないとされていた27)。筆者らは、従来法では可視化できない領域ごとの移動制御が存在するのではと仮説を立てた。脳室にフルオレセイン(蛍光色素の一種)の前駆体を注入したところ、注入後数時間以内の間に脳室面全体でM期を迎える細胞をラベルできた。大脳皮質発生最初期(胎生10日)から最後期(胎生17日)までの各ステージに産生されるニューロンをラベルし経時的に追跡し記載した28)。これまで記載されていない種々の興味深い現象が可視化されたが、とくに胎生12–15日ラベルのニューロンは背内側のほうが背外側よりも早く脳表面に到達することがわかった(図1B)。これは、詳細な組織学的解析やライブイメージングにより、移動途中に通過する背外側サブプレート下での滞留によると考えられた。ここには視床皮質投射線維束も横走しているが、ゲノム編集により視床に発現するGbx2をノックアウトすることで視床皮質投射線維をなくしたとき29, 30)にも移動の領域差は観察されたが、サブプレートが脳表面に誤配置される変異マウス(reeler)では領域差が小さくなった。このことから、背外側サブプレートという早生まれニューロンの構造が、後に移動するニューロンの移動を制御する可能性が示唆された。なお、視床皮質投射線維はニューロン産生と分化を制御していることを示唆する報告がその後なされた31)。大脳皮質には種々の領野があるが、移動制御の違いによって回路形成や機能が異なってくれば面白い。
この論文では、ニューロン産生最初期から、後期まで、高い時空間分解能によるラベルと追跡を行い、大脳皮質発生研究者の本棚には必ず入っている古典的な教科書27)(トリチウムチミジンをつかって、増殖と移動を追跡)をアップデートする包括的な記載を行った。またこの方法は、脳全体で、特定の誕生時刻のニューロンを高い時間分解能で簡便にラベルできるメリットがあり、筆者らも前障ニューロンの移動の記述などに活用した例を報告しているほか32)、ノックアウトマウスや薬剤投与モデルでの移動フェノタイプの検出に優位性がある。ご興味のある方はご一読・ご活用いただけると幸いである。
このような組織における細胞レベルでの制御を解析する過程で、意図せず「メタ細胞群」としての理解すべき移動・配置異常を観察したので、まとめて報告した33)。発生期においては移動細胞(群)が周囲の細胞(群)(これらはいずれも同じ神経幹細胞群に由来する)と相互作用しながら細胞秩序を形成するので、生態学における個体を細胞に置き換えたメタファーが成立すると考えられる。メタ細胞群とは、生態学におけるメタ個体群に概念に対応する概念として筆者が考えた造語である(不均一環境での生物の移動は、生物群集の中で間接的に相互作用する個体群におおきな影響を与えるが、不均一な環境で構成種の移動によって繋がる局所個体群の集合をメタ個体群と呼ぶ)。
先に軽く触れたように、大脳皮質発生において、inside-outパターンが逆転する現象が、Reelinやその下流のDab1を欠損した34–39)動物で観察されてきた。この場合、早生まれのニューロンは遅生まれのニューロンを追い越せない。一方、Dab1を欠損した表層ニューロンはサブプレートを越えて皮質板に移動できること40, 41)、Dab1は細胞質または核に存在して細胞自律的に機能する42–46)ことが過去の報告で知られていたが、それではReelin/Dab1経路を欠損したマウスでなぜ層構造が逆転するのかは説明できなかった。背理法的にDab1の機能には、広義の細胞非自律的機能があると仮説を立てた。まず筆者らはDab1 floxマウスで表層ニューロンが産生される胎生14.5日にEPを行い、Dab1-/-表層ニューロン(CreによってloxPに挟まれた配列が切り出されてノックアウトされた細胞を簡単のためDab1-/-と表現する)は皮質板に進入できることを再確認した(図1C)。また、Dab1 floxマウスで深層ニューロンが産生される胎生12.5日にEPを行い、Dab1-/-深層ニューロンは皮質板に入れないことも再確認した。胎生12.5日にEPを行ったDab1 floxマウスで表層ニューロンマーカーを染色したところ、皮質板には入れない(=サブプレート下に誤配置されている)深層ニューロンの密度が高い脳においてのみ、Dab1-/-表層ニューロンがサブプレート下に誤集積していた。Creレポーターマウスを用いた実験により、誤集積したニューロンの大半は、胎生12.5日にEPされた神経幹細胞の娘細胞の子孫であることを確認した。この表現型は前向きにノックアウト効率を下げたり、Dab1-/-深層ニューロン特異的にDab1を強制発現させたりすることで救済された。すなわち、密なDab1-/-深層ニューロン群はDab1-/-浅層ニューロンの皮質板への進入を妨げた。これはDab1の細胞自律的ではない機能の反映と考えられ、これまで長年謎だった、Reelin/Dab1経路を欠損した哺乳類でなぜ層構造が逆転するのかという問いに対して、一定の答えを得られた。また、別の言い方をすれば、一部の細胞群の移動・配置異常が、連鎖的にその後同じ場所を通過すべき細胞群の配置異常を連鎖的に引き起こすことがわかり、Reelin/Dab1経路はこうしたメタ細胞群としての配置異常を防ぐ機能があることがわかった。
この研究では、
——を駆使することで、メタ細胞群としての配置異常を実験的に検証できた。これまで細胞自律的異常や、細胞とその細胞外環境の相互作用の異常はこれまで多くの研究者が調べてきたが、移動異常には、細胞レベルを超えた異常があることを提示したことには、一定の意義があると考えている。白質ニューロンの数的以上は精神疾患死後脳で繰り返し報告されており、これまでは古典的染色や正常サルでの実験に基づきこれがサブプレートの遺残だと考えられることが多いが47, 48)が、精神神経疾患脳においては小児神経疾患でみられるようなシビアな移動異常が一部の細胞のみで生じて白質ニューロンの増加等がおきている可能性も提起している。最近は、精神疾患における発達障害関連遺伝子の体細胞モザイク変異といった考え方もでてきているが49)、関連した現象を観察している可能性もあり、詳細な神経病理学的解析が要請される。
統合失調症をはじめとした精神神経疾患の生物学的研究は盛んに行われている。臨床的な神経画像・生理学的研究は高度に数学的になり、遺伝学的研究も急速に解像度と包括性を増し、動物モデルは着実に所見を積み重ね、疾患iPS細胞やオルガノイドの研究も進んでいるが、細胞・分子レベルの病態がなかなか見えてこない。再現性ある病理学的所見に乏しいため「神経病理学者の墓場」と言われている状況は本質的には変わりがない。筆者は近年勃興した死後脳のオミクス研究に興味をもっているが、その分子方向の解像度、空間解像度、サンプル数、空間情報のデータが増していくはずである。こうしたあらゆる視点での精神神経疾患研究を横糸でつなぐのは、神経解剖学、神経生理学といった、きわめて基礎的な神経生物学だろうと考えている。ここに時間軸を付け加える神経発生学の重要性はますます高まっていくと感じている。
本稿で紹介した筆者の研究は、慶應義塾大学医学部解剖学教室、東京慈恵会医科大学解剖学講座での研究で得られた知見である。研究遂行にあたり、多大なご指導を賜った仲嶋一範先生、久保健一郎先生、田畑秀典先生、仲嶋研究室・久保研究室の皆様、共同研究者の先生がた、ならびに技術員の方々に感謝申し上げる。また、本稿で紹介した研究内容は、日本学術振興会、文部科学省、日本医療研究開発機構、上原記念生命科学財団、先進医薬研究振興財団、武田科学財団、慶應義塾学事振興基金、福澤諭吉記念慶應義塾学事振興基金、慶應義塾大学医学部研究奨励費、慶應医師会医学研究助成金、慈恵大学萌芽的共同研究推進費からの研究費により行われた。最後に、執筆機会を与えて下さった日本神経化学会出版・広報委員会、奨励賞選考委員会の皆様に御礼申し上げる。
1) Takahashi T, Nowakowski RS, Caviness VS Jr. The leaving or Q fraction of the murine cerebral proliferative epithelium: a general model of neocortical neuronogenesis. J Neurosci, 16(19), 6183–6196 (1996).
2) Angevine JB, Bodian D, Coulombre AJ, Edds MV, Hamburger V, Jacobson M, Lyser KM, Prestige MC, Sidman RL, Varon S, Weiss PA; Boulder-Committee. Embryonic vertebrate central nervous system: revised terminology. The Boulder Committee. Anat Rec, 166(2), 257–261 (1970).
3) Tabata H, Kanatani S, Nakajima K. Differences of migratory behavior between direct progeny of apical progenitors and basal progenitors in the developing cerebral cortex. Cereb Cortex, 19(9), 2092–2105 (2009).
4) Sekine K, Honda T, Kawauchi T, Kubo K, Nakajima K. The outermost region of the developing cortical plate is crucial for both the switch of the radial migration mode and the Dab1-dependent “inside-out” lamination in the neocortex. J Neurosci, 31(25), 9426–9439 (2011).
5) Shin M, Kitazawa A, Yoshinaga S, Hayashi K, Hirata Y, Dehay C, Kubo KI, Nakajima K. Both excitatory and inhibitory neurons transiently form clusters at the outermost region of the developing mammalian cerebral neocortex. J Comp Neurol, 527(10), 1577–1597 (2019).
6) Yozu M, Tabata H, Nakajima K. Birth-date dependent alignment of GABAergic neurons occurs in a different pattern from that of non-GABAergic neurons in the developing mouse visual cortex. Neurosci Res, 49(4), 395–403 (2004).
7) Stoner R, Chow ML, Boyle MP, Sunkin SM, Mouton PR, Roy S, Wynshaw-Boris A, Colamarino SA, Lein ES, Courchesne E. Patches of disorganization in the neocortex of children with autism. N Engl J Med, 370(13), 1209–1219 (2014).
8) Stevens JR, Casanova MF. Is there a neuropathology of schizophrenia? Biol Psychiatry, 24(2), 123–128 (1988).
9) Kubo KI. Increased densities of white matter neurons as a cross-disease feature of neuropsychiatric disorders. Psychiatry Clin Neurosci, 74(3), 166–175 (2020).
10) Yoshinaga S, Ohkubo T, Sasaki S, Nuriya M, Ogawa Y, Yasui M, Tabata H, Nakajima K. A Phosphatidylinositol Lipids System, Lamellipodin, and Ena/VASP Regulate Dynamic Morphology of Multipolar Migrating Cells in the Developing Cerebral Cortex. J Neurosci, 32(34), 11643–11656 (2012).
11) Tabata H, Nakajima K. Multipolar migration: the third mode of radial neuronal migration in the developing cerebral cortex. J Neurosci, 23(31), 9996–10001 (2003).
12) Noctor SC, Martinez-Cerdeno V, Ivic L, Kriegstein AR. Cortical neurons arise in symmetric and asymmetric division zones and migrate through specific phases. Nat Neurosci, 7(2), 136–144 (2004).
13) Stensaas LJ, Stensaas SS. An electron microscope study of cells in the matrix and intermediate laminae of the cerebral hemisphere of the 45 mm rabbit embryo. Z Zellforsch Mikrosk Anat, 91(3), 341–365 (1968).
14) Hatanaka Y, Yamauchi K. Excitatory cortical neurons with multipolar shape establish neuronal polarity by forming a tangentially oriented axon in the intermediate zone. Cereb Cortex, 23(1), 105–113 (2013).
15) Namba T, Kibe Y, Funahashi Y, Nakamuta S, Takano T, Ueno T, Shimada A, Kozawa S, Okamoto M, Shimoda Y, Oda K, Wada Y, Masuda T, Sakakibara A, Igarashi M, Miyata T, Faivre-Sarrailh C, Takeuchi K, Kaibuchi K. Pioneering axons regulate neuronal polarization in the developing cerebral cortex. Neuron, 81(4), 814–829 (2014).
16) Rakic P. Mode of cell migration to the superficial layers of fetal monkey neocortex. J Comp Neurol, 145(1), 61–83 (1972).
17) LoTurco JJ, Bai J. The multipolar stage and disruptions in neuronal migration. Trends Neurosci, 29(7), 407–413 (2006).
18) Kawauchi T, Chihama K, Nabeshima Y, Hoshino M. Cdk5 phosphorylates and stabilizes p27kip1 contributing to actin organization and cortical neuronal migration. Nat Cell Biol, 8(1), 17–26 (2006).
19) Krause M, Leslie JD, Stewart M, Lafuente EM, Valderrama F, Jagannathan R, Strasser GA, Rubinson DA, Liu H, Way M, Yaffe MB, Boussiotis VA, Gertler FB. Lamellipodin, an Ena/VASP ligand, is implicated in the regulation of lamellipodial dynamics. Dev Cell, 7(4), 571–583 (2004).
20) Michael M, Vehlow A, Navarro C, Krause M. c-Abl, Lamellipodin, and Ena/VASP proteins cooperate in dorsal ruffling of fibroblasts and axonal morphogenesis. Curr Biol, 20(9), 783–791 (2010).
21) Bae YH, Ding Z, Das T, Wells A, Gertler F, Roy P. Profilin1 regulates PI(3,4)P2 and lamellipodin accumulation at the leading edge thus influencing motility of MDA-MB-231 cells. Proc Natl Acad Sci USA, 107(50), 21547–21552 (2010).
22) Chang C, Adler CE, Krause M, Clark SG, Gertler FB, Tessier-Lavigne M, Bargmann CI. MIG-10/lamellipodin and AGE-1/PI3K promote axon guidance and outgrowth in response to slit and netrin. Curr Biol, 16(9), 854–862 (2006).
23) Pinheiro EM, Xie Z, Norovich AL, Vidaki M, Tsai LH, Gertler FB. Lpd depletion reveals that SRF specifies radial versus tangential migration of pyramidal neurons. Nat Cell Biol, 13(8), 989–995 (2011).
24) Caviness VS Jr., Sidman RL. Time of origin of corresponding cell classes in the cerebral cortex of normal and reeler mutant mice: an autoradiographic analysis. J Comp Neurol, 148(2), 141–151 (1973).
25) Caviness VS Jr. Neocortical histogenesis in normal and reeler mice: a developmental study based upon [3H]thymidine autoradiography. Brain Res, 256(3), 293–302 (1982).
26) Ogawa M, Miyata T, Nakajimat K, Yagyu K, Seike M, Ikenaka K, Yamamoto H, Mikoshibat K. The reeler gene-associated antigen on cajal-retzius neurons is a crucial molecule for laminar organization of cortical neurons. Neuron, 14(5), 899–912 (1995).
27) Bayer SA. Altman J. Neocortical development, (Raven Press New York, 1991).
28) Yoshinaga S, Shin M, Kitazawa A, Ishii K, Tanuma M, Kasai A, Hashimoto H, Kubo KI, Nakajima K. Comprehensive characterization of migration profiles of murine cerebral cortical neurons during development using FlashTag labeling. iScience, 24(4), 102277 (2021).
29) Hevner RF, Miyashita-Lin E, Rubenstein JL. Cortical and thalamic axon pathfinding defects in Tbr1, Gbx2, and Pax6 mutant mice: evidence that cortical and thalamic axons interact and guide each other. J Comp Neurol, 447(1), 8–17 (2002).
30) Miyashita-Lin EM, Hevner R, Wassarman KM, Martinez S, Rubenstein JL. Early neocortical regionalization in the absence of thalamic innervation. Science, 285(5429), 906–909 (1999).
31) Monko T, Rebertus J, Stolley J, Salton SR, Nakagawa Y. Thalamocortical axons regulate neurogenesis and laminar fates in the early sensory cortex. Proc Natl Acad Sci USA, 119(22), e2201355119 (2022).
32) Oshima K, Yoshinaga S, Kitazawa A, Hirota Y, Nakajima K, Kubo KIA. Unique “reversed” migration of neurons in the developing claustrum. J Neurosci, 43(5), 693–708 (2023).
33) Yoshinaga S, Honda T, Kubo KI, Nakajima K. Dab1-deficient deep layer neurons prevent Dab1-deficient superficial layer neurons from entering the cortical plate. Neurosci Res, 180, 23–35 (2022).
34) González JL, Russo CJ, Goldowitz D, Sweet HO, Davisson MT, Walsh CA. Birthdate and cell marker analysis of scrambler: a novel mutation affecting cortical development with a reeler-like phenotype. J Neurosci, 17(23), 9204–9211 (1997).
35) Howell BW, Gertler FB, Cooper JA. Mouse disabled (mDab1): a Src binding protein implicated in neuronal development. EMBO J, 16(1), 121–132 (1997).
36) Kojima T, Nakajima K, Mikoshiba K. The disabled 1 gene is disrupted by a replacement with L1 fragment in yotari mice. Brain Res Mol Brain Res, 75(1), 121–127 (2000).
37) Rice DS, Sheldon M, D’Arcangelo G, Nakajima K, Goldowitz D, Curran T. Disabled-1 acts downstream of Reelin in a signaling pathway that controls laminar organization in the mammalian brain. Development, 125(18), 3719–3729 (1998).
38) Sheldon M, Rice DS, D’Arcangelo G, Yoneshima H, Nakajima K, Mikoshiba K, Howell BW, Cooper JA, Goldowitz D, Curran T. Scrambler and yotari disrupt the disabled gene and produce a reeler -like phenotype in mice. Nature, 389(6652), 730–733 (1997).
39) Yoneshima H, Nagata E, Matsumoto M, Yamada M, Nakajima K, Miyata T, Ogawa M, Mikoshiba K. A novel neurological mutant mouse, yotari, which exhibits reeler-like phenotype but expresses CR-50 antigen/Reelin. Neurosci Res, 29(3), 217–223 (1997).
40) Franco SJ, Martinez-Garay I, Gil-Sanz C, Harkins-Perry SR, Muller U. Reelin regulates cadherin function via Dab1/Rap1 to control neuronal migration and lamination in the neocortex. Neuron, 69(3), 482–497 (2011).
41) Sekine K, Kawauchi T, Kubo K, Honda T, Herz J, Hattori M, Kinashi T, Nakajima K. Reelin controls neuronal positioning by promoting cell-matrix adhesion via inside-out activation of integrin alpha5beta1. Neuron, 76(2), 353–369 (2012).
42) Honda T, Nakajima K. Mouse Disabled1 (DAB1) is a nucleocytoplasmic shuttling protein. J Biol Chem, 281(50), 38951–38965 (2006).
43) Honda T, Nakajima K. Proper level of cytosolic disabled-1, which is regulated by dual nuclear translocation pathways, is important for cortical neuronal migration. Cereb Cortex, 26(7), 3219–3236 (2016).
44) Hammond V, Howell B, Godinho L, Tan SS. Disabled-1 functions cell autonomously during radial migration and cortical layering of pyramidal neurons. J Neurosci, 21(22), 8798–8808 (2001).
45) Morimura T, Ogawa M. Relative importance of the tyrosine phosphorylation sites of Disabled-1 to the transmission of Reelin signaling. Brain Res, 1304, 26–37 (2009).
46) Sanada K, Gupta A, Tsai LH. Disabled-1-regulated adhesion of migrating neurons to radial glial fiber contributes to neuronal positioning during early corticogenesis. Neuron, 42(2), 197–211 (2004).
47) Kostovic I, Judas M, Sedmak G. Developmental history of the subplate zone, subplate neurons and interstitial white matter neurons: relevance for schizophrenia. Int J Dev Neurosci, 29(3), 193–205 (2011).
48) Kostovic I, Rakic P. Developmental history of the transient subplate zone in the visual and somatosensory cortex of the macaque monkey and human brain. J Comp Neurol, 297(3), 441–470 (1990).
49) Nishioka M, Takayama J, Sakai N, Kazuno AA, Ishiwata M, Ueda J, Hayama T, Fujii K, Someya T, Kuriyama S, Tamiya G, Takata A, Kato T. Deep exome sequencing identifies enrichment of deleterious mosaic variants in neurodevelopmental disorder genes and mitochondrial tRNA regions in bipolar disorder. Mol Psychiatry, (2023).

This page was created on 2023-11-07T09:58:50.326+09:00
This page was last modified on 2024-01-26T13:11:58.000+09:00
このサイトは(株)国際文献社によって運用されています。