中枢神経系の発達、障害を司る脳内免疫システム
国立精神・神経医療研究センター 神経研究所 神経薬理研究部
発行日:2024年6月30日Published: June 30, 2024
© 2024 日本神経化学会© 2024 The Japanese Society for Neurochemistry
従来、中枢神経系には末梢の免疫系細胞は存在せず、リンパ管もないなどの理由から中枢神経系は免疫系が介在しない免疫寛容な臓器として考えられてきた歴史がある1)。しかし、近年の研究からその常識は覆され、中枢神経系の特定の領域には免疫系細胞が存在し、脳の発達や恒常性の維持に寄与するほか、神経変性疾患や中枢性の自己免疫疾患の際には中枢神経系に浸潤して神経系の障害過程に関わるなど免疫系細胞が中枢神経系の発達、機能、障害を司る「脳内免疫システム」が形成されていることが明らかになってきた。しかし、脳内免疫システムを構成する免疫系細胞は発達段階や病態などによって種類や局在、遺伝子発現パターンが大きく変化することから、中枢神経系の発達や障害過程における免疫系細胞の役割は多岐にわたる。本稿では、中枢神経系の発達や障害を制御する脳内免疫システムの役割とそのメカニズムについて最新の知見を交えて概説する。
脳に存在する免疫系細胞は、脳の実質に常在するミクログリアとそれ以外の特定の領域にしか存在しない末梢由来の免疫系細胞に大別される。ミクログリアは卵黄嚢に由来するミエロイド系の免疫系細胞であり、胎生期に脳に移行して中枢神経系の各所に常在するようになり、中枢神経系の生体防御や神経回路の形成に関わる2)。一方、T細胞やB細胞といったリンパ球やマクロファージ、好中球などのミエロイド系細胞は生理条件下では血液脳関門を透過できないために脳の実質にはほとんど存在せず、髄膜や脈絡叢、血管周囲腔といった末梢と中枢の境界領域に局在する(図1左)。脳機能に対する影響は解明されていないことが多いが、T細胞の欠損マウスでは成体神経新生が抑制されること3)、認知機能の低下が観察されることからT細胞は生理的な脳機能を制御する重要な因子であることが伺える4)。生理条件下では境界領域に局在して脳機能を制御している免疫系細胞であるが、脳神経疾患の病態下では全く異なる性質を示す。多発性硬化症や視神経脊髄炎といった炎症性脳神経疾患、筋萎縮性側索硬化症やアルツハイマー病といった神経変性疾患では血液脳関門の透過性が減弱し、血中の免疫系細胞が中枢神経系の実質に浸潤して炎症を引き起こす(図1右)5)。これらの免疫系細胞は神経やグリア細胞に作用して神経変性や脱髄にも寄与する。このように免疫系細胞は生理条件下と病態下では全く異なる顔を持ち、脳の発達や障害過程に深く関わる。下記では、筆者がこれまで手掛けてきた脳内免疫システムによる脳の発達制御や障害のメカニズムについて詳述する。
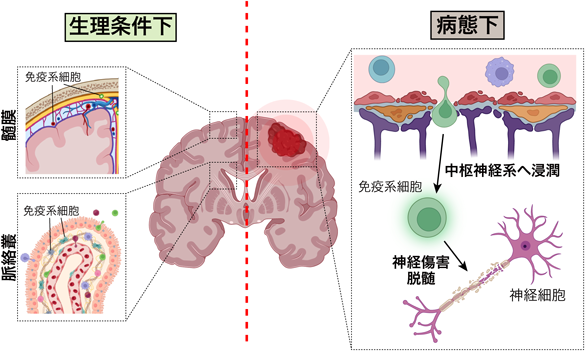
生理条件下では髄膜や脈絡叢といった中枢と末梢の境界領域に局在し、成体神経新生や生体防御に働く。病態下では、血液脳関門が減弱して免疫系細胞が中枢神経系に浸潤して炎症を惹起するとともに神経傷害や脱髄を引き起こす。
末梢に由来する免疫系細胞は海馬の成体神経新生やミエリンの再形成など神経系の細胞に作用して脳機能を制御する3, 6)。また、ミクログリアは免疫系のシグナルを通して軸索の保護、シナプスの形成、剪定など脳の発達過程に強く影響する7)。このように、免疫系は神経細胞やグリア細胞に作用して脳の発生、発達を支える重要な因子であるが、末梢由来の免疫系細胞が直接的に脳の発達過程に寄与するのかは不明であった。そこで、著者らははじめに発達段階の脳に存在する免疫系細胞の種類と量をフローサイトメトリーで解析した。その結果、生後1日から5日程度の新生児期のマウスの脳ではB細胞が最も豊富に存在するなど成体とは全く異なるポピュレーションを示すことを発見した。B細胞はその役割によってサブタイプが分けられる。抗原提示によって特定の抗体を産生するB-2細胞、特異性の低い自然抗体を産生して自然免疫に関わるB-1aおよびB-1b細胞、炎症を抑制させる制御性B細胞などがある8)。新生児期のマウスに局在するB細胞のサブタイプを特定するためにフローサイトメトリーで細胞表面マーカーを解析したところ、発達期の脳におけるB細胞はCD5+ CD43+ CD1d+ CD19+ CD45R+のB-1a細胞であることを見出した。ここで疑問となってくることが、「B細胞がどのようなメカニズムで脳へ移行するのか」ということと、「B-1a細胞は脳の発達に対して何らかの機能を有するのか」という点である。1つ目の疑問である遊走のメカニズムを明らかにするため、B細胞に作用するケモカインの同定を試みた。B細胞はCXCL12-CXCR4やCXCL13-CXCR5といったケモカインのシグナルを通じて組織に遊走される9, 10)。新生児期の血中におけるB細胞のケモカイン受容体の発現を解析したところCXCR5を強く発現しており、そのリガンドであるCXCL13は脈絡叢に強く発現することが確認された。さらに、CXCL13に対する機能阻害抗体を胎生期のマウスの脳室内に投与すると新生児期の脳に局在するB細胞が著しく減少した。このことから、B細胞はCXCL13-CXCR5のシグナルを通じて脳へ移行することが示唆される(図2)。
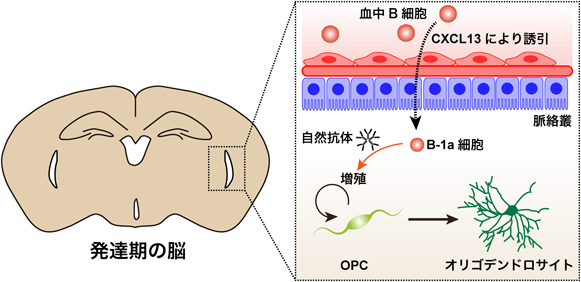
発達期の脳において、B細胞は脈絡叢から産生されるCXCL13により血中から誘引され、脈絡叢や髄膜などの境界領域に局在する。このB細胞はB-1a細胞へと性質を変化させ、自然抗体を産生してOPCの増殖を促し、オリゴデンドロサイトの成熟化を制御する。
2つ目の疑問点であるB-1a細胞による脳の発達に対する機能を明らかにするため、B細胞を除去することのできるBAFFR抗体を胎生期のマウスの脳室内に投与した。その結果、新生児期の白質においてミエリンを形成する細胞であるオリゴデンドロサイトの数が減少していたとともに、オリゴデンドロサイトの前駆細胞であるOPC(Oligodendrocyte precursor cell)の増殖が抑制されていることが確認された。この結果から、B-1a細胞はOPCの増殖を促進することでオリゴデンドロサイトの成熟化に寄与していることが示唆された。次に、B-1a細胞がOPCの増殖を促進する分子メカニズムを探索した。B-1a細胞は自然抗体を産生して生体防御に関わる細胞である。また、OPCは自然抗体に対するFc受容体であるFcα/μRを発現している。そこで、B-1a細胞から産生される自然抗体がOPCの増殖を促進しているのかを検討するために、Fcα/μRに対する機能阻害抗体を投与した。その結果、増殖性のOPCやオリゴデンドロサイトの数が減少し、ミエリンを巻いている軸索の数も減少することが確認された。さらに、自然抗体を新生児期のマウスの脳に投与すると増殖性のOPCやオリゴデンドロサイトの数が増加した。これらの結果から、B-1a細胞は自然抗体を産生することで発達期のOPCの増殖を促進し、ミエリンの形成に寄与することを明らかにした(図2)11)。
中枢神経系の自己免疫疾患である多発性硬化症では、末梢の免疫系細胞が中枢神経系に浸潤して炎症を惹起し、脱髄や軸索変性を誘導することで四肢の麻痺や視覚障害などの神経症状を引き起こす。特に、炎症を誘導する主要な免疫系細胞であるCD4+ T細胞は炎症性サイトカインの産生や神経との直接的な接触により神経系を傷害する12)。しかし、T細胞が神経系を障害する具体的な分子メカニズムは十分明らかにはなっていなかった。そんな中、筆者らは免疫系細胞が多発性硬化症の病態を悪化させる因子としてRepulsive guidance molecule-a(RGMa)という分子に着目していた。RGMaは軸索の伸長を阻害する機能を持つGlycosylphosphatidylinositol(GPI)アンカー型の膜タンパク質であるが、それ以外にも細胞死や免疫応答を制御するなど多彩な機能を持つ分子である13)。免疫系細胞が神経系を障害するメカニズムにRGMaが関わる可能性を考え、CD4+ T細胞におけるRGMaの発現を解析した。CD4+ T細胞はTh1, Th17, 制御性T細胞など性質によってサブタイプが分かれるが、その中でも炎症を強く引き起こすT細胞であるTh17細胞がRGMaを強く発現していることを見出した。Th17細胞は炎症を誘導する以外にも、神経と直接的に接触することで傷害性を発揮することがin vivo imagingの研究で示されていた12)。RGMaは膜タンパク質であるため、Th17細胞はRGMaを通して神経と接触して神経傷害を引き起こしている可能性を着想した。そこで、RGMaの機能阻害抗体の存在下でTh17細胞と神経細胞を共培養したところ、Th17細胞により誘導される神経傷害がRGMa機能阻害抗体により抑制された。さらに、Th17細胞が発現するRGMaがin vivoでも神経傷害性を持つのかを検証するため、中枢神経系の抗原を認識するTh17細胞をマウスに移植することでTh17細胞を原因とする脳脊髄炎のモデルを作製し、RGMaの機能阻害抗体を投与した14)。RGMa抗体投与群では、神経症状の進行が阻害され、脊髄で観察される軸索の変性が妨げられた。さらに、RGMaが軸索を変性させるシグナル経路をTh17細胞と神経細胞の共培養実験で探索したところ、RGMa抗体の存在下では対照抗体を添加した群と比べてリン酸化したAktが多く検出された。これらの結果から、Th17細胞はRGMaを通して神経細胞のAktを脱リン酸化し、神経傷害を引き起こすことが示唆された(図3)15)。
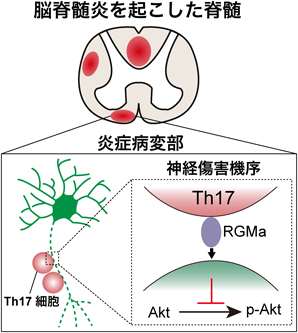
脳脊髄炎が起こると免疫系細胞が中枢神経系に浸潤して炎症を引き起こす。この炎症が生じている病変部ではTh17細胞がRGMaというタンパク質を通して神経のAktを脱リン酸化させ、神経を傷害する。
多発性硬化症患者の症状は典型的には再発と寛解を繰り返す特徴があるが、一部の患者では次第に再発がない進行性の経過を辿る二次進行型多発性硬化症に病型が変化する16)。再発寛解型の多発性硬化症に対しては免疫系細胞を標的にした薬剤がいくつか開発されているものの、二次進行型多発性硬化症に対する治療薬は乏しい。RGMaは神経傷害や炎症など作用点を複数持つことから、RGMaの機能阻害抗体が二次進行型多発性硬化症に対しても治療効果を持つ可能性を検討した。そこで、二次進行型多発性硬化症のモデルマウスを作製し、RGMaの機能阻害抗体を投与する実験を行った。二次進行型多発性硬化症モデルマウスとして、1型糖尿病の自然発症マウスであるnon-obese diabetic(NOD)マウスに対してミエリンの構成タンパク質を抗原として免疫するExperimental Autoimmune Encephalomyelitis(EAE)を誘導するモデルを作製した17)。このマウスでは、一過性に強い神経症状が観察された後に一度寛解し、その後に進行性の症状を示す二次進行型に移行する。このモデル動物の二次進行期にRGMa抗体を投与したところ、神経症状の進行が抑制され、脊髄における炎症や軸索の変性が減弱することも確認された18)。このことから、RGMa抗体は二次進行性の脳脊髄炎を抑制させる作用を持つことが示唆され、RGMa抗体が二次進行型多発性硬化症の病態に対して有効な治療法となることが期待される。
中枢神経系が免疫寛容な臓器と考えられていた時代は終わり、免疫系細胞は中枢神経系の発生、恒常性維持、障害に深く関わることがわかってきたほか、これまでの常識を覆す知見も多く報告されていることから神経免疫学は近年飛躍的に発展してきた研究分野であるといえる。歴史上、脳には存在しないと考えられていたリンパ管も発見されたほか19)、single cell RNA-seqなどの1細胞解析技術の発展で老化や神経変性疾患の脳における免疫系細胞のサブポピュレーションや遺伝子発現パターンが続々と明らかになってきている20)。しかし、脳内免疫システムによる脳の発達や脳機能の制御機構、脳神経疾患の病態形成上の機能をさらに詳細に理解していくためには、脳内免疫システムや脳内の細胞群はもちろん血管やリンパ管などの脈管系なども含めて中枢神経系を包括的に捉えていく必要がある。今後、脳内免疫システムが脳機能を支えるさらなるメカニズムや病態における機能が解明され、脳神経疾患の治療標的となる細胞、分子が発見されていくことが期待される。
本稿で紹介させていただきました研究は、大阪大学大学院医学系研究科で実施いたしました。本研究に関しまして、多くのご支援とご指導を賜りました大阪大学大学院医学系研究科 山下俊英教授および研究室の皆様、共同研究者の先生方にこの場を借りて深く感謝申し上げます。また、この度執筆の機会を与えていただきました神経化学会出版・広報委員会の澤本和延委員長、委員の先生方ならびに関係者の皆様に深く御礼申し上げます。
1) Carson MJ, Doose JM, Melchior B, Schmid CD, Ploix CC. CNS immune privilege: hiding in plain sight. Immunol Rev, 213(1), 48–65 (2006).
2) Ginhoux F, Greter M, Leboeuf M, Nandi S, See P, Gokhan S, Mehler MF, Conway SJ, Ng LG, Stanley ER, Samokhvalov IM, Merad M. Fate mapping analysis reveals that adult microglia derive from primitive macrophages. Science, 330(6005), 841–845 (2010).
3) Ziv Y, Ron N, Butovsky O, Landa G, Sudai E, Greenberg N, Cohen H, Kipnis J, Schwartz M. Immune cells contribute to the maintenance of neurogenesis and spatial learning abilities in adulthood. Nat Neurosci, 9(2), 268–275 (2006).
4) Derecki NC, Cardani AN, Yang CH, Quinnies KM, Crihfield A, Lynch KR, Kipnis J. Regulation of learning and memory by meningeal immunity: A key role for IL-4. J Exp Med, 207(5), 1067–1080 (2010).
5) Yong VW, Rivest S. Taking advantage of the systemic immune system to cure brain diseases. Neuron, 64(1), 55–60 (2009).
6) Durak O, Gao F, Kaeser-Woo YJ, Rueda R, Martorell AJ, Nott A, Liu CY, Watson LA, Tsai LH. Chd8 mediates cortical neurogenesis via transcriptional regulation of cell cycle and Wnt signaling. Nat Neurosci, 19(11), 1477–1488 (2016).
7) Butovsky O, Weiner HL. Microglial signatures and their role in health and disease. Nat Rev Neurosci, 19(10), 622–635 (2018).
8) Baumgarth N. The double life of a B-1 cell: Self-reactivity selects for protective effector functions. Nat Rev Immunol, 11(1), 34–46 (2011).
9) Bleul CC, Schultze JL, Springer TA. B Lymphocyte chemotaxis regulated in association with microanatomic localization, differentiation state, and B cell receptor engagement. J Exp Med, 187(5), 753–762 (1998).
10) Ansel KM, Harris RBS, Cyster JG. CXCL13 is required for B1 cell homing, natural antibody production, and body cavity immunity. Immunity, 16(1), 67–76 (2002).
11) Tanabe S, Yamashita T. B-1a lymphocytes promote oligodendrogenesis during brain development. Nat Neurosci, 21(4), 506–516 (2018).
12) Siffrin V, Radbruch H, Glumm R, Niesner R, Paterka M, Herz J, Leuenberger T, Lehmann SM, Luenstedt S, Rinnenthal JL, Laube G, Luche H, Lehnardt S, Fehling HJ, Griesbeck O, Zipp F. In vivo imaging of partially reversible th17 cell-induced neuronal dysfunction in the course of encephalomyelitis. Immunity, 33(3), 424–436 (2010).
13) Siebold C, Yamashita T, Monnier PP, Mueller BK, Pasterkamp RJ. RGMs: Structural insights, molecular regulation, and downstream signaling. Trends Cell Biol, 27(5), 365–378 (2017).
14) Jäger A, Dardalhon V, Sobel RA, Bettelli E, Kuchroo VK. Th1, Th17, and Th9 effector cells induce experimental autoimmune encephalomyelitis with different pathological phenotypes. J Immunol, 183(11), 7169–7177 (2009).
15) Tanabe S, Yamashita T. Repulsive guidance molecule-a is involved in Th17-Cell-induced neurodegeneration in autoimmune encephalomyelitis. Cell Rep, 9(4), 1459–1470 (2014).
16) Reich DS, Lucchinetti CF, Calabresi PA. Multiple sclerosis. N Engl J Med, 378(2), 169–180 (2018).
17) Basso AS, Frenkel D, Quintana FJ, Costa-Pinto FA, Petrovic-Stojkovic S, Puckett L, Monsonego A, Bar-Shir A, Engel Y, Gozin M, Weiner HL. Reversal of axonal loss and disability in a mouse model of progressive multiple sclerosis. J Clin Invest, 118(4), 1532–1543 (2008).
18) Tanabe S, Fujita Y, Ikuma K, Yamashita T. Inhibiting repulsive guidance molecule-a suppresses secondary progression in mouse models of multiple sclerosis. Cell Death Dis, 9(11), 1–10 (2018).
19) Louveau A, Smirnov I, Keyes TJ, Eccles JD, Rouhani SJ, Peske JD, Derecki NC, Castle D, Mandell JW, Lee KS, Harris TH, Kipnis J. Structural and functional features of central nervous system lymphatic vessels. Nature, 523(7560), 337–341 (2015).
20) Gate D, Saligrama N, Leventhal O, Yang AC, Unger MS, Middeldorp J, Chen K, Lehallier B, Channappa D, De Los Santos MB, McBride A, Pluvinage J, Elahi F, Tam GK, Kim Y, Greicius M, Wagner AD, Aigner L, Galasko DR, Davis MM, Wyss-Coray T. Clonally expanded CD8 T cells patrol the cerebrospinal fluid in Alzheimer’s disease. Nature, 577(7790), 399–404 (2020).

This page was created on 2024-06-12T11:02:35.536+09:00
This page was last modified on 2024-06-17T15:01:41.000+09:00
このサイトは(株)国際文献社によって運用されています。