アストロサイトによる血液脳関門機能制御
金沢大学医薬保健研究域医学系神経解剖学
発行日:2019年6月30日Published: June 30, 2019
© 2019 日本神経化学会© 2019 The Japanese Society for Neurochemistry
脳は外界から隔離された組織である。頭蓋骨内で髄膜中に満たされた脳脊髄液に浮かび物理的な衝撃から守られているだけでなく、目に見えないレベルにおいてもバリア構造を備え、他の臓器と異なり毒素や病原体から守られている。その実態をなすシステムが血液脳関門(blood-brain barrier: BBB)であり、血中と脳組織間の物質移動が厳密にコントロールされる1, 2)。この物理的、機能的なバリアが存在することで、脳はその機能を正しく発揮することができる。近年、脳梗塞や脳損傷、神経変性疾患など異なる神経性疾患の病態において、BBBの機能破綻が関与することがわかってきた3)。研究技術の発展もあり、BBBの機能破綻に関する知見がこの20年余りで飛躍的に蓄積されてきている。本稿では、BBBの機能とその破綻について概説するとともに、筆者らが最近報告した脳虚血後のBBB破綻のアストロサイトによる制御機構4)について紹介したい。
BBBは血液と脳組織間の物質移動を制限しているバリアシステムであり、中枢神経系の微小環境の恒常性を維持している。脳内の血管は、タイトジャンクション(tight junction: TJ)を持つ血管内皮細胞、ペリサイト、細胞外基質から構成され、さらにアストロサイトの終足に取り囲まれた構造を持つ。近年はこれら細胞に神経細胞やミクログリアを含めた、Neurovascular unitという概念が提唱されている(図1)。脳血管細胞と脳実質細胞が密接に関わり、多細胞間コミュニケーションにより、BBB機能や神経活動依存の脳血流、血管新生や神経炎症が調節されることが明らかとなりつつある5, 6)。バリアとしての血中と脳組織の物質移動制御において、TJを持つ血管内皮細胞がその機能主体と考えられる。脳内の血管内皮細胞は、脳室周囲器官や脈絡叢など一部を除き、細胞間隙の透過を制御するClaudin, OccludinなどのTJタンパク質を発現している。これにより受動輸送は阻止されるとともに、P-gp(P-glycoprotein)等の輸送体や受容体が能動的な排出あるいは供給を担い、脳に必要な分子と不要な分子の通過を調節している。しかしBBBの物理的バリアとしての特性は、血管内皮細胞のみにより形成されるわけではない。血管内皮細胞の周囲に存在するぺリサイトは、積極的にBBBの形成、血流調節、免疫細胞トラフィッキングに関わることが、PDGF-B(platelet-derived growth factor-B)またはその受容体PDGFRβの欠損によるペリサイト欠失マウスの解析から報告されている7, 8)。他の末梢臓器血管と異なり、中枢神経系血管ではペリサイトが豊富であることもその機能の重要性を反映していると考えられる。一方、アストロサイトから産生されるShh(sonic hedgehog)は、Shh欠損マウスおよび下流分子Smo(smoothened)欠損マウスの解析から、血管が形成されたあとのBBBの機能成熟に寄与することが報告されている9)。また、アストロサイトから分泌されるApoE(apolipoprotein E)は、アルツハイマー病の重要なリスク因子であり、ApoE欠損マウスやヒトAPOEノックインマウスの解析からペリサイトの機能調節を介してBBB透過性を制御することが示されている10)。
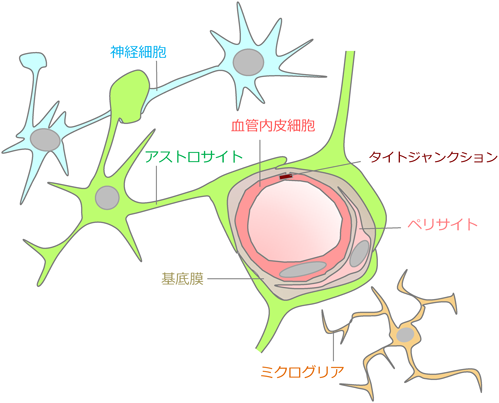
生理的条件下において脳と血中の物質移動は血液脳関門により厳密に制御される。脳毛細血管はタイトジャンクションを持つ血管内皮細胞、ペリサイト、細胞外基質により構成され、アストロサイトの終足に取り囲まれる。さらに神経細胞やミクログリアなどを含む機能単位における細胞間相互作用により脳血管の機能が調節される。
BBBの機能分子であるTJが失われると、脳実質への免疫細胞の浸潤や分子の流入がおこり、恒常性の破綻と神経炎症を引き起こす。酸化ストレスや炎症などによりBBBの機能破綻が誘導されるが、この現象は脳血管疾患である脳梗塞だけでなく、神経変性疾患である筋萎縮性側索硬化症や脱髄疾患の多発性硬化症でも観察される。また、代表的な神経変性疾患の1つであるアルツハイマー病においては、初期血管ダメージからのBBBの機能障害がAβのクリアランス低下と産生亢進につながり認知機能を低下させる、という血管仮説が提唱されている11, 12)。このように一部の疾患ではBBBの機能破綻は結果ではなく、病態形成の原因としても注目されはじめており、神経細胞の障害とBBBの機能破綻には密接な関係性が示唆される。その中でも、臨床上BBBの機能制御が重要な意味を持つ疾患として脳梗塞が挙げられる。
脳梗塞では、致死をまぬがれても寝たきり等の重篤な後遺症を引き起こす。現在唯一有効な薬理学的治療はt-PA(tissue-plasminogen activator)による血栓溶解療法であるが、その適用はより重篤な出血性脳梗塞をきたすリスクのため脳梗塞発症後4.5時間以内の患者に限られる13)。この出血性脳梗塞への転換は、BBBの機能破綻に起因し治療可能時間枠拡大の枷となっている。脳虚血後のBBB破綻は、末梢免疫細胞の脳内浸潤や、その後の炎症の遷延、予後の悪化と密接に関係することが明らかとなっている。脳虚血後の神経障害の拡大を抑えるため、末梢細胞と脳実質細胞の相互作用およびBBBの調節機構を明らかにすることは治療戦略上重要な意義を持つ。これまで、BBBの機能制御解析は血管内皮細胞やペリサイトなど血管細胞機能に重点がおかれ、アストロサイトの役割には未だ不明な点が多い。今回我々は、アストロサイトに発現する分子NDRG2(N-myc downstream-regulated gene 2)に着目し、脳虚血後のアストロサイトのBBB破綻への関与を明らかにした。
NDRG2は中枢神経系においてアストロサイト特異的に発現する分子である。腫瘍抑制因子としての作用が知られ、その発現は低酸素をはじめステロイドホルモンや重金属など種々のストレスにより上昇する14, 15)。我々はこれまでにNDRG2がアストロサイトの活性化に促進的に働くことを脳損傷モデルで明らかにしているが16)、ラット一過性脳虚血モデルにおいてもNDRG2の発現が上昇することが報告されている17)。そこでNDRG2を介したアストロサイトの脳梗塞病態への関与を検証した。脳虚血モデルは、一過性脳虚血と比して脳梗塞巣サイズが安定し、長期経過が観察可能な、永久中大脳動脈閉塞モデルを用いた。免疫組織化学の結果から、NDRG2は脳虚血後1日という早期から、虚血周辺部において発現上昇することが明らかになった。NDRG2欠損マウスを用いた解析では、脳虚血2週間後における脳梗塞巣サイズの拡大が認められ、内在性のアストロサイトNDRG2が保護的な役割を担う可能性が示唆された。同マウスでは虚血周辺部のGFAP陽性活性化アストロサイトによるGlial scarが低形成であり、Iba1陽性ミクログリア/マクロファージの集積は亢進していた。さらに、フローサイトメトリーによる解析から、虚血半球におけるマクロファージ、T細胞、B細胞の脳内浸潤が亢進していることが示唆された。しかし脳虚血後、アストロサイト以外の脳実質細胞や脳内に浸潤してきたCD45陽性細胞にはNDRG2の発現は認められなかった。これらの結果からアストロサイトのNDRG2が白血球の脳内浸潤を制御することで脳梗塞病態に保護的に働く可能性が示唆された。
アストロサイトのNDRG2はどのように白血球の脳内浸潤を制御するのだろうか。白血球の脳内浸潤に細胞特異性は認められなかったことから、脳血管の強度に違いがあるのではないかと仮説を立てた。また、NDRG2の発現上昇の応答が早期であることから脳梗塞後早期に起こる現象への関与が考えられ、BBBの機能破綻に注目した。脳梗塞後には、数時間から数日の時間幅でBBBの破綻が引き起こされる18)。静脈よりトレーサーを投与し、脳内におけるトレーサーの血管外漏出を組織学的に検出したところ、脳虚血領域におけるシグナルがNDRG2欠損マウスで亢進していた。さらに、ウエスタンブロットおよび免疫組織化学解析から、内在性血中タンパク質についても、虚血側における高発現および血管周囲への漏出の亢進がNDRG2欠失により認められた。以上の結果から、アストロサイトは脳梗塞後にNDRG2の発現を増加させ、血管透過性を制御することが示唆された。
BBBの強度を制御するメカニズムを明らかにするために、DNAマイクロアレイを用いて網羅的な解析を行った。その結果、脳虚血後NDRG2欠損により発現が上昇した遺伝子の上位にMMP(matrix metalloproteinase)ファミリーの1つMMP-3が認められた。MMPファミリーは細胞外基質の分解に関与し、組織リモデリングやがんの転移に重要な役割を担う。脳梗塞におけるBBB破綻や出血において、MMP-2およびMMP-9が主要な役割を担うことが、遺伝子欠損マウスおよび阻害薬の解析から明らかとなっている19–21)。また、MMP-3も欠損マウスの解析から、t-PA投与脳虚血モデルでの出血に寄与すること、脊髄損傷およびパーキンソン病モデルのBBB破綻に寄与することが報告されている22–24)。定量的PCR法を用いて、NDRG2欠失による脳虚血後のMMP発現への関与を検証したところ、上記3種MMPのうちMMP-3およびMMP-9の発現上昇の有意な亢進が認められた。そこで免疫組織化学によりMMP-3およびMMP-9の組織内分布を検討したところ、MMP-9は主に虚血中心部のGr-1陽性好中球に存在しており、MMP-3は虚血周辺部のS100β陽性NDRG2陽性アストロサイトに高発現していた。MMP-9の虚血側における酵素活性、およびMMP-9陽性Gr-1陽性好中球数はNDRG2の欠損により有意に増加した。これらの結果は、NDRG2が細胞自律性にMMP-3を、非細胞自律性にMMP-9を制御する可能性を示唆している。そこで、NDRG2によってMMP-3が真に細胞自律性に制御されるかを培養アストロサイトで検証した。その結果NDRG2欠損アストロサイトでは、細胞内MMP-3発現およびIL-1β誘導性に分泌された細胞外MMP-3発現が有意に増加した。これにより、アストロサイト内のNDRG2がMMP-3の発現を規定することが明らかとなった。これまでの解析からNDRG2の脳虚血病態への関与として2つのメカニズムが考えられる。一つは脳虚血後早期のMMP-3の発現抑制によって、BBBの機能破綻とそれに続く炎症性細胞の脳内浸潤を抑制する働きである。他方は、Glial scar形成を促進することで虚血領域周囲に壁をつくり、正常領域への炎症の拡大を制限する働きである。脳梗塞後にアストロサイトで発現上昇するNDRG2はこの2種の制御機構により脳虚血後の組織障害に保護的な役割を担うと示唆される(図2)。
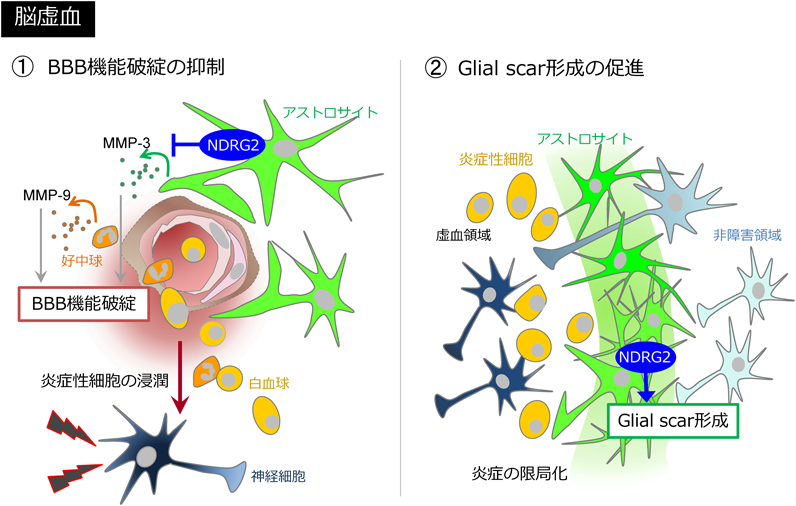
脳虚血後にアストロサイトで発現が上昇するNDRG2は、MMPの発現制御を介して虚血早期に起こる血液脳関門の破綻を抑制し、続く白血球浸潤を抑制する。その後Glial scarの形成を促進することで、非障害領域への炎症の拡大を抑制する。NDRG2はこの2つの作用を介し脳虚血病態に保護作用を示す。(文献4より改変)
著者らは上記のように、脳梗塞病態においてアストロサイトのNDRG2がMMP-3を介しBBB機能破綻を制御することを明らかにした。NDRG2はこれまで、その発現の低さとヒトにおけるがんの悪性度・転移・予後の悪さとの相関が知られていたが、がん研究領域だけではなく、BBB機能破綻を伴う神経性疾患領域においても病態に関与する可能性が示唆される。今後、出血性脳梗塞とNDRG2発現レベルの相関など、脳梗塞患者における知見の蓄積が期待される。
アストロサイトは、古典的に病態下ではグリア瘢痕を形成し神経軸索再生を阻害すると考えられていたが、近年にはアストロサイトが神経傷害後の組織修復や軸索再生に寄与することも見出されている25)。そして、様々な病態下で活性化するアストロサイトは遺伝子発現を劇的に変えるが、その病態に応じて炎症性のA1アストロサイト、抗炎症性でA2アストロサイトと異なる特性を示すという概念も提唱されている26)。遺伝子発現プロファイリングにより、LPS投与後のアストロサイト(A1)と比較し、脳虚血後のアストロサイト(A2)は神経保護因子を豊富に発現するなど異なる遺伝子特性を持つことが示された27)。また脳虚血後には、アストロサイトのCD38を介した神経細胞へのミトコンドリア転移が起こり、神経症状の回復に寄与するという現象も報告されている28)。一方で、脳虚血慢性期に抑制性T細胞が活性化アストロサイトを抑制することで脳機能修復に寄与することや29)、脳梗塞後にアストロサイトが局所脳血流を低下させること30)、アストロサイト活性化の減弱に関連し脳血流の低下や神経症状が緩和されることも報告されている31)。免疫細胞から脳を守るBBB機能や、神経細胞のエネルギー需要に応じた脳血流の供給など、病態下で失われる血管恒常性を取り戻すことが、神経機能の回復に重要となると考えられる。
神経性疾患の病態におけるアストロサイトの多様な働きへの関心はこの数年益々高まってきている。均一かと思われていたアストロサイトの集団や応答は、その部位や取り巻く病態の状況によって大きく異なることがわかってきた。アストロサイト機能の理解のためにグリアの不均一性と部位特性を明らかにすることが鍵となり、同時に異なるアストロサイト集団に共通して高発現する分子の機能も重要となる32)。NDRG2は後者の観点からアストロサイトとの本質に迫る糸口になりうると考える。グリア細胞機能への介入が神経疾患の治療法開発の突破口となるよう、グリアと血管、神経、免疫細胞間における、多細胞間コミュニケーションの詳細のさらなる解明が期待される。
本稿で紹介した研究の遂行にあたり、ご指導を賜りました金沢大学医薬保健研究域医学系神経解剖学講座の堀修教授、ならびに研究室の皆様、共同研究者の先生方に、心より感謝申し上げます。本研究内容は、日本学術振興会、文部科学省、内藤科学技術振興財団からの研究費により行われました。最後に、このような執筆の機会を与えて下さいました出版・広報員会の澤本和延教授ならびに神経化学編集部の皆様に厚く御礼申し上げます。
1) Abbott NJ, Patabendige AA, Dolman DE, Yusof SR, Begley DJ. Structure and function of the blood-brain barrier. Neurobiol Dis, 37(1), 13–25 (2010).
2) Zlokovic BV. The blood-brain barrier in health and chronic neurodegenerative disorders. Neuron, 57(2), 178–201 (2008).
3) Sweeney MD, Sagare AP, Zlokovic BV. Blood-brain barrier breakdown in Alzheimer disease and other neurodegenerative disorders. Nat Rev Neurol, 14(3), 133–150 (2018).
4) Takarada-Iemata M, Yoshikawa A, Ta HM, Okitani N, Nishiuchi T, Aida Y, Kamide T, Hattori T, Ishii H, Tamatani T, Le TM, Roboon J, Kitao Y, Matsuyama T, Nakada M, Hori O. N-myc downstream-regulated gene 2 protects blood-brain barrier integrity following cerebral ischemia. Glia, 66(7), 1432–1446 (2018).
5) Obermeier B, Daneman R, Ransohoff RM. Development, maintenance and disruption of the blood-brain barrier. Nat Med, 19(12), 1584–1596 (2013).
6) Iadecola C. The Neurovascular Unit Coming of Age: A Journey through Neurovascular Coupling in Health and Disease. Neuron, 96(1), 17–42 (2017).
7) Armulik A, Genove G, Mae M, Nisancioglu MH, Wallgard E, Niaudet C, He L, Norlin J, Lindblom P, Strittmatter K, Johansson BR, Betsholtz C. Pericytes regulate the blood-brain barrier. Nature, 468(7323), 557–561 (2010).
8) Daneman R, Zhou L, Kebede AA, Barres BA. Pericytes are required for blood-brain barrier integrity during embryogenesis. Nature, 468(7323), 562–566 (2010).
9) Alvarez JI, Dodelet-Devillers A, Kebir H, Ifergan I, Fabre PJ, Terouz S, Sabbagh M, Wosik K, Bourbonniere L, Bernard M, van Horssen J, de Vries HE, Charron F, Prat A. The Hedgehog pathway promotes blood-brain barrier integrity and CNS immune quiescence. Science, 334(6063), 1727–1731 (2011).
10) Bell RD, Winkler EA, Singh I, Sagare AP, Deane R, Wu Z, Holtzman DM, Betsholtz C, Armulik A, Sallstrom J, Berk BC, Zlokovic BV. Apolipoprotein E controls cerebrovascular integrity via cyclophilin A. Nature, 485(7399), 512–516 (2012).
11) Kisler K, Nelson AR, Montagne A, Zlokovic BV. Cerebral blood flow regulation and neurovascular dysfunction in Alzheimer disease. Nat Rev Neurosci, 18(7), 419–434 (2017).
12) Nation DA, Sweeney MD, Montagne A, Sagare AP, D’Orazio LM, Pachicano M, Sepehrband F, Nelson AR, Buennagel DP, Harrington MG, Benzinger TLS, Fagan AM, Ringman JM, Schneider LS, Morris JC, Chui HC, Law M, Toga AW, Zlokovic BV. Blood-brain barrier breakdown is an early biomarker of human cognitive dysfunction. Nat Med, 25(2), 270–276 (2019).
13) Hacke W, Kaste M, Bluhmki E, Brozman M, Davalos A, Guidetti D, Larrue V, Lees KR, Medeghri Z, Machnig T, Schneider D, von Kummer R, Wahlgren N, Toni D. ECASS Investigators. Thrombolysis with alteplase 3 to 4.5 hours after acute ischemic stroke. N Engl J Med, 359(13), 1317–1329 (2008).
14) Hu W, Fan C, Jiang P, Ma Z, Yan X, Di S, Jiang S, Li T, Cheng Y, Yang Y. Emerging role of N-myc downstream-regulated gene 2 (NDRG2) in cancer. Oncotarget, 7(1), 209–223 (2016).
15) Qu X, Zhai Y, Wei H, Zhang C, Xing G, Yu Y, He F. Characterization and expression of three novel differentiation-related genes belong to the human NDRG gene family. Mol Cell Biochem, 229(1–2), 35–44 (2002).
16) Takarada-Iemata M, Kezuka D, Takeichi T, Ikawa M, Hattori T, Kitao Y, Hori O. Deletion of N-myc downstream-regulated gene 2 attenuates reactive astrogliosis and inflammatory response in a mouse model of cortical stab injury. J Neurochem, 130(3), 374–387 (2014).
17) Li Y, Shen L, Cai L, Wang Q, Hou W, Wang F, Zeng Y, Zhao G, Yao L, Xiong L. Spatial-temporal expression of NDRG2 in rat brain after focal cerebral ischemia and reperfusion. Brain Res, 1382, 252–258 (2011).
18) Yang Y, Rosenberg GA. Blood-brain barrier breakdown in acute and chronic cerebrovascular disease. Stroke, 42(11), 3323–3328 (2011).
19) Asahi M, Wang X, Mori T, Sumii T, Jung JC, Moskowitz MA, Fini ME, Lo EH. Effects of matrix metalloproteinase-9 gene knock-out on the proteolysis of blood-brain barrier and white matter components after cerebral ischemia. J Neurosci, 21(19), 7724–7732 (2001).
20) Liu J, Jin X, Liu KJ, Liu W. Matrix metalloproteinase-2-mediated occludin degradation and caveolin-1-mediated claudin-5 redistribution contribute to blood-brain barrier damage in early ischemic stroke stage. J Neurosci, 32(9), 3044–3057 (2012).
21) Svedin P, Hagberg H, Savman K, Zhu C, Mallard C. Matrix metalloproteinase-9 gene knock-out protects the immature brain after cerebral hypoxia-ischemia. J Neurosci, 27(7), 1511–1518 (2007).
22) Chung YC, Kim YS, Bok E, Yune TY, Maeng S, Jin BK. MMP-3 contributes to nigrostriatal dopaminergic neuronal loss, BBB damage, and neuroinflammation in an MPTP mouse model of Parkinson’s disease. Mediators Inflamm, 2013, 370526 (2013).
23) Lee JY, Choi HY, Ahn HJ, Ju BG, Yune TY. Matrix metalloproteinase-3 promotes early blood-spinal cord barrier disruption and hemorrhage and impairs long-term neurological recovery after spinal cord injury. Am J Pathol, 184(11), 2985–3000 (2014).
24) Suzuki Y, Nagai N, Umemura K, Collen D, Lijnen HR. Stromelysin-1 (MMP-3) is critical for intracranial bleeding after t-PA treatment of stroke in mice. J Thromb Haemost, 5(8), 1732–1739 (2007).
25) Anderson MA, Burda JE, Ren Y, Ao Y, O’Shea TM, Kawaguchi R, Coppola G, Khakh BS, Deming TJ, Sofroniew MV. Astrocyte scar formation aids central nervous system axon regeneration. Nature, 532(7598), 195–200 (2016).
26) Liddelow SA, Barres BA. Reactive Astrocytes: Production, Function, and Therapeutic Potential. Immunity, 46(6), 957–967 (2017).
27) Zamanian JL, Xu L, Foo LC, Nouri N, Zhou L, Giffard RG, Barres BA. Genomic analysis of reactive astrogliosis. J Neurosci, 32(18), 6391–6410 (2012).
28) Hayakawa K, Esposito E, Wang X, Terasaki Y, Liu Y, Xing C, Ji X, Lo EH. Transfer of mitochondria from astrocytes to neurons after stroke. Nature, 535(7613), 551–555 (2016).
29) Ito M, Komai K, Mise-Omata S, Iizuka-Koga M, Noguchi Y, Kondo T, Sakai R, Matsuo K, Nakayama T, Yoshie O, Nakatsukasa H, Chikuma S, Shichita T, Yoshimura A. Brain regulatory T cells suppress astrogliosis and potentiate neurological recovery. Nature, 565(7738), 246–250 (2019).
30) Chuquet J, Hollender L, Nimchinsky EA. High-resolution in vivo imaging of the neurovascular unit during spreading depression. J Neurosci, 27(15), 4036–4044 (2007).
31) Begum G, Song S, Wang S, Zhao H, Bhuiyan MIH, Li E, Nepomuceno R, Ye Q, Sun M, Calderon MJ, Stolz DB, St Croix C, Watkins SC, Chen Y, He P, Shull GE, Sun D. Selective knockout of astrocytic Na+ /H+ exchanger isoform 1 reduces astrogliosis, BBB damage, infarction, and improves neurological function after ischemic stroke. Glia, 66(1), 126–144 (2018).
32) Chai H, Diaz-Castro B, Shigetomi E, Monte E, Octeau JC, Yu X, Cohn W, Rajendran PS, Vondriska TM, Whitelegge JP, Coppola G, Khakh BS. Neural Circuit-Specialized Astrocytes: Transcriptomic, Proteomic, Morphological, and Functional Evidence. Neuron, 95(3), 531–549.e539 (2017).

This page was created on 2019-05-07T10:41:33.337+09:00
This page was last modified on 2019-07-03T11:25:32.000+09:00
このサイトは(株)国際文献社によって運用されています。