活性酸素種感受性TRPチャネルによる血管性認知障害の病態制御機構の解明
和歌山県立医科大学 薬学部 病院薬学研究室
発行日:2025年12月30日Published: December 30, 2025
© 2024 日本神経化学会© 2024 The Japanese Society for Neurochemistry
超高齢社会の現代において認知症患者は増加の一途をたどっており、社会的な課題となっている。一方で、認知症の新規治療薬の創出は難航しており既存の治療薬の満足度も未だ低いままである。近年、軽度認知障害からアルツハイマー病や血管性認知症を含む多くの認知症に共通して脳血管障害が認められることが知られ始め1)、脳血管障害が認知症の主要な原因の1つであることが支持されるようになってきた。脳血管障害に起因する認知機能障害として血管性認知障害(vascular cognitive impairment: VCI)という疾患概念が提唱され、VCIを惹起する主な要因として加齢や生活習慣病などに伴い脳血流の穏やかな低下が続く慢性脳低灌流状態が知られている。慢性脳低灌流状態を伴う多くの動物モデルや認知症患者で神経細胞傷害に先立つ髄鞘の傷害や脱落(白質傷害)が観察され、VCIの特徴的で重要な病変として注目されてきた2, 3)。しかし、慢性脳低灌流状態により惹起されるVCIや白質傷害の病態制御機構には不明な点が多く、認知症の根本的治療法開発が難航している一因となっている。
慢性脳低灌流状態により惹起されるVCIの研究には、マウスの両側総頸動脈に内径0.18 mmのマイクロコイルを装着する手術により慢性脳低灌流状態を惹起する両側総頸動脈狭窄(Bilateral common carotid artery stenosis: BCAS)モデルマウスが多く用いられてきた(図1)4, 5)。BCASモデルマウスはVCIの臨床像をよく反映し、神経細胞傷害に先立ち手術の約4週後から認知機能障害と白質傷害が認められることが知られている。さらにBCASモデルマウスではグリア細胞の活性化が認められ、活性酸素種(reactive oxygen species: ROS)が病態に関与することが示唆されている2, 6)。
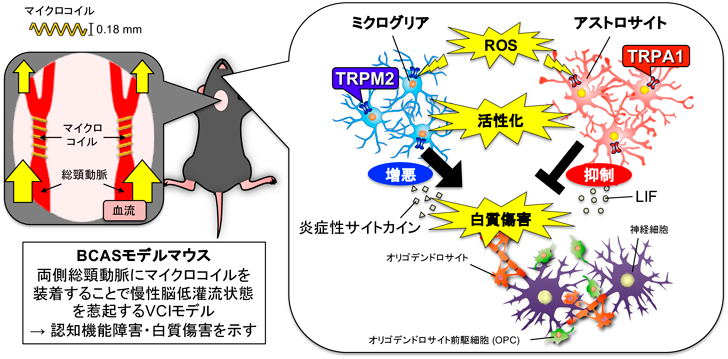
そこで筆者は、慢性脳低灌流状態に伴い発症する認知機能障害、白質傷害の病態制御機構の解明を目指し、グリア細胞に機能的に発現するROS感受性transient receptor potential(TRP)チャネルのTRPM2およびTRPA1に着目して研究を進めてきた。本稿ではVCI病態におけるTRPM2とミクログリア、TRPA1とアストロサイトの病態制御機構についての筆者らの研究成果7–9)を紹介する。
TRPM2は脳内のミクログリア10)、神経細胞11)や単球・マクロファージ12)、好中球13)などの末梢免疫細胞などに機能的に発現するROS感受性のCa2+透過性非選択的カチオンチャネルである。TRPM2は炎症性サイトカイン/ケモカインなどの産生を介した様々な炎症性疾患への関与が報告されている14)。一方で筆者らは、食餌投与によりミクログリアを除去できるCSF1R阻害薬のPLX3397を用いて、慢性脳低灌流によって惹起されるVCIにおいてミクログリアを介した脳内炎症が病態増悪に関与することを見出した15)。そこで本稿では、TRPM2とミクログリアに着目し慢性脳低灌流や加齢に伴い認められる認知機能障害、白質傷害への脳内炎症の関与について研究を行った。
BCASモデルマウスを用いた解析により、BCAS処置28日後において野生型マウスで認知機能障害、白質傷害が認められたのに対し、TRPM2欠損マウスでは抑制された。VCIにおけるミクログリアの機能へのTRPM2の関与を調べるため、白質が豊富な脳部位の脳梁において免疫染色を行ったところ、BCAS処置により野生型マウスではIBA1陽性ミクログリア数が増加したがTRPM2欠損マウスでは抑制された。またBCAS処置28日後の脳梁において、野生型マウスではIL1βやTNFαなどの炎症性サイトカインが増加したがTRPM2欠損マウスでは抑制された。
次に加齢に伴う認知機能低下へのTRPM2、ミクログリアの関与を調べるため高齢マウスを用いた解析を行った。20–24ヶ月齢の高齢野生型マウスでは認知機能低下、白質傷害が認められたのに対し、高齢TRPM2欠損マウスでは認められなかった。また脳梁において高齢野生型マウスで認められたIBA1陽性ミクログリア数やTnfa発現の増加が高齢TRPM2欠損マウスでは認められなかった。
以上より、慢性脳低灌流や加齢に伴う認知機能障害、白質傷害の病態増悪にTRPM2とミクログリアの活性化に伴う脳内炎症が関与していることを明らかにした(図1)7, 8)。
TRPA1は最もredox感受性の高いTRPチャネルで16)、知覚神経の侵害受容器に発現し様々な痛みに関与することが知られている17)。中枢神経系において、TRPA1はアストロサイト18–20)、血管内皮細胞21)、オリゴデンドロサイト22)などに発現し、アルツハイマー病や脳血管疾患への関与が報告されている22–25)。本稿では慢性脳低灌流により惹起されるVCIにおけるTRPA1の役割に着目し、脳内に最も豊富に存在するグリア細胞であるアストロサイトとの関与についてBCASモデルマウスを用いて検証した。
野生型マウスにおいて、BCAS処置により惹起される認知機能障害、白質傷害は、モデル作製28日後の後期で観察されたが、14日後の早期では観察されなかった。一方で、TRPA1欠損マウスでは、BCAS処置14日後の早期から認知機能障害、白質傷害が認められた。次に、BCAS処置後の野生型マウスにTRPA1刺激薬のシンナムアルデヒドを連日投与することで、BCAS処置28日後の後期に認められた認知機能障害、白質傷害が抑制された。VCIにおけるアストロサイトの機能へのTRPA1の関与を調べるため、免疫染色や定量的PCRを用いた組織学的解析を行ったところ、野生型マウスでBCAS処置14日後の早期から脳梁でのGFAP陽性アストロサイト数やGfap遺伝子発現が増加したが、TRPA1欠損マウスでは増加しなかった。そこでCre-loxPシステムを用いて細胞種特異的TRPA1欠損マウスを作製し検討を行ったところ、全身TRPA1欠損マウスと同様にBCAS処置14日後の早期において、アストロサイト特異的TRPA1欠損マウスでも認知機能障害、白質傷害が認められた。一方、TRPA1の発現が知られる血管内皮細胞やオリゴデンドロサイト系譜細胞の特異的TRPA1欠損マウスでは認知機能障害、白質傷害は観察されなかった。これより、アストロサイトのTRPA1がVCIにおいて病態抑制の役割を担っている可能性が示された。
次にTRPA1によるVCI病態抑制の分子機序を明らかにするため、RNAseqにより遺伝子発現変動を網羅的に解析した。BCAS処置によりTRPA1欠損マウスと比べ野生型マウスでアストロサイト関連遺伝子の発現増加が認められた。発現変動が認められた遺伝子についてGene Ontologyエンリッチメント解析を行い、上位に見出されたアストロサイト関連遺伝子の中から髄鞘化促進作用のある白血病阻止因子(leukemia inhibitory factor: LIF)26)に着目し解析を行った。脳梁やMACSにより分取したアストロサイトにおける定量的PCRにより、BCAS処置14日後の野生型マウスではLifの発現増加が認められたのに対し、TRPA1欠損マウスでは遺伝子発現に変化は認められなかった。さらに脳梁の免疫染色により、BCAS処置14日後の野生型マウスではGFAP陽性アストロサイトでLIFの増加が認められたのに対し、TRPA1欠損マウスでは認められなかった。そこで、慢性脳低灌流により惹起されるVCIへのLIFの作用を評価するため、LIF受容体を構成するgp130の阻害薬のSC144や抗LIF中和抗体を野生型マウスに投与し解析を行った。BCAS処置14日後の早期の野生型マウスでは認められなかった認知機能障害、白質傷害がSC144の連日腹腔内投与や抗LIF中和抗体の脳室内投与により観察された。これよりTRPA1を介したアストロサイトからのLIFの増加が病態抑制に関与することが示された。
TRPA1を介した病態抑制機序をさらに詳細に明らかにするため、初代培養細胞を用いた実験を行った。初代培養アストロサイトにTRPA1刺激作用のあるH2O2を処置すると、野生型アストロサイトでPKC/p38の活性化を介したLifの発現増加が認められた。さらにH2O2を処置した野生型アストロサイトの培養上清を初代培養オリゴデンドロサイト前駆細胞(OPC)に適用することで髄鞘を形成する成熟オリゴデンドロサイトへの分化が促進された。一方で、TRPA1欠損アストロサイトではH2O2処置によるLifの発現増加やアストロサイト培養上清によるOPCの成熟オリゴデンドロサイトへの分化促進作用は認められなかった。またH2O2を処置した野生型アストロサイトの培養上清に抗LIF中和抗体を処置することで、OPCに対する分化促進作用が抑制された。
以上より、アストロサイトのTRPA1活性化がLIFの産生を介して髄鞘形成を促進することで、VCIにおける内因性の生体防御機構を担っていることを明らかにした(図1)9)。
最近、アルツハイマー病において新たに疾患修飾薬が承認され注目を集めているが、解決すべき課題も多く残されており、認知症の新規治療薬の創出は未だに難航している。近年、アルツハイマー病を含む多くの認知症をVCIとして捉え、認知症の主要な原因として脳血管障害やそれに伴う白質傷害の重要性が支持されるものの、脳血管障害や白質傷害を対象に新規治療標的を探索する研究は十分に進んでいない。脳血流の低下や白質傷害はうつ病などの他の中枢神経疾患においても観察されることが知られており27)、治療対象としての白質傷害の重要性を明らかにした本稿の研究成果は、VCIを始めとした多くの中枢神経疾患に対する創薬標的探索のための有望な研究戦略となることが期待される。
本稿で紹介した研究成果は、京都大学大学院薬学研究科生体機能解析学分野で得られたものです。本研究の遂行にあたり多大なるご指導を賜った金子周司先生、白川久志先生、中川貴之先生、永安一樹先生をはじめとした研究室の方々、京都大学大学院工学研究科の森泰生先生、山梨大学大学院総合研究部医学域の小泉修一先生、繁冨英治先生をはじめとした共同研究者の先生方に、心より感謝申し上げます。また、この度、本稿の執筆機会を与えて下さいました日本神経化学会奨励賞選考委員の先生方、ならびに編集部、関係者の皆様に深く感謝申し上げます。
1) Iadecola C. The neurovascular unit coming of age: A journey through neurovascular coupling in health and disease. Neuron, 96(1), 17–42 (2017).
2) Saggu R, Schumacher T, Gerich F, Rakers C, Tai K, Delekate A, Petzold GC. Astroglial NF-kB contributes to white matter damage and cognitive impairment in a mouse model of vascular dementia. Acta Neuropathol Commun, 4(1), 76 (2016).
3) Dichgans M, Leys D. Vascular cognitive impairment. Circ Res, 120(3), 573–591 (2017).
4) Shibata M, Ohtani R, Ihara M, Tomimoto H. White matter lesions and glial activation in a novel mouse model of chronic cerebral hypoperfusion. Stroke, 35(11), 2598–2603 (2004).
5) Kakae M, Kawashita A, Onogi H, Nakagawa T, Shirakawa H. Bilateral common carotid artery stenosis in mice: A model of chronic cerebral hypoperfusion-induced vascular cognitive impairment. Bio Protoc, 14(13), e5022 (2024).
6) Miyamoto N, Maki T, Pham LD, Hayakawa K, Seo JH, Mandeville ET, Mandeville JB, Kim KW, Lo EH, Arai K. Oxidative stress interferes with white matter renewal after prolonged cerebral hypoperfusion in mice. Stroke, 44(12), 3516–3521 (2013).
7) Miyanohara J, Kakae M, Nagayasu K, Nakagawa T, Mori Y, Arai K, Shirakawa H, Kaneko S. TRPM2 channel aggravates CNS inflammation and cognitive impairment via activation of microglia in chronic cerebral hypoperfusion. J Neurosci, 38(14), 3520–3533 (2018).
8) Kakae M, Miyanohara J, Morishima M, Nagayasu K, Mori Y, Shirakawa H, Kaneko S. Pathophysiological role of TRPM2 in age-related cognitive impairment in mice. Neuroscience, 408, 204–213 (2019).
9) Kakae M, Nakajima H, Tobori S, Kawashita A, Miyanohara J, Morishima M, Nagayasu K, Nakagawa T, Shigetomi E, Koizumi S, Mori Y, Kaneko S, Shirakawa H. The astrocytic TRPA1 channel mediates an intrinsic protective response to vascular cognitive impairment via LIF production. Sci Adv, 9(29), eadh0102 (2023).
10) Miyake T, Shirakawa H, Kusano A, Sakimoto S, Konno M, Nakagawa T, Mori Y, Kaneko S. TRPM2 contributes to LPS/IFNγ-induced production of nitric oxide via the p38/JNK pathway in microglia. Biochem Biophys Res Commun, 444(2), 212–217 (2014).
11) Kaneko S, Kawakami S, Hara Y, Wakamori M, Itoh E, Minami T, Takada Y, Kume T, Katsuki H, Mori Y, Akaike A. A critical role of TRPM2 in neuronal cell death by hydrogen peroxide. J Pharmacol Sci, 101(1), 66–76 (2006).
12) Yamamoto S, Shimizu S, Kiyonaka S, Takahashi N, Wajima T, Hara Y, Negoro T, Hiroi T, Kiuchi Y, Okada T, Kaneko S, Lange I, Fleig A, Penner R, Nishi M, Takeshima H, Mori Y. TRPM2-mediated Ca2+influx induces chemokine production in monocytes that aggravates inflammatory neutrophil infiltration. Nat Med, 14(7), 738–747 (2008).
13) Hiroi T, Wajima T, Negoro T, Ishii M, Nakano Y, Kiuchi Y, Mori Y, Shimizu S. Neutrophil TRPM2 channels are implicated in the exacerbation of myocardial ischaemia/reperfusion injury. Cardiovasc Res, 97(2), 271–281 (2013).
14) Haraguchi K, Kawamoto A, Isami K, Maeda S, Kusano A, Asakura K, Shirakawa H, Mori Y, Nakagawa T, Kaneko S. TRPM2 contributes to inflammatory and neuropathic pain through the aggravation of pronociceptive inflammatory responses in mice. J Neurosci, 32(11), 3931–3941 (2012).
15) Kakae M, Tobori S, Morishima M, Nagayasu K, Shirakawa H, Kaneko S. Depletion of microglia ameliorates white matter injury and cognitive impairment in a mouse chronic cerebral hypoperfusion model. Biochem Biophys Res Commun, 514(4), 1040–1044 (2019).
16) Takahashi N, Kuwaki T, Kiyonaka S, Numata T, Kozai D, Mizuno Y, Yamamoto S, Naito S, Knevels E, Carmeliet P, Oga T, Kaneko S, Suga S, Nokami T, Yoshida J, Mori Y. TRPA1 underlies a sensing mechanism for O2. Nat Chem Biol, 7(10), 701–711 (2011).
17) Bautista DM, Pellegrino M, Tsunozaki M. TRPA1: A gatekeeper for inflammation. Annu Rev Physiol, 75(1), 181–200 (2013).
18) Shigetomi E, Tong X, Kwan KY, Corey DP, Khakh BS. TRPA1 channels regulate astrocyte resting calcium and inhibitory synapse efficacy through GAT-3. Nat Neurosci, 15(1), 70–80 (2011).
19) Shigetomi E, Jackson-Weaver O, Huckstepp RT, O’Dell TJ, Khakh BS. TRPA1 channels are regulators of astrocyte basal calcium levels and long-term potentiation via constitutive D-serine release. J Neurosci, 33(24), 10143–10153 (2013).
20) Oh SJ, Lee JM, Kim HB, Lee J, Han S, Bae JY, Hong GS, Koh W, Kwon J, Hwang ES, Woo DH, Youn I, Cho IJ, Bae YC, Lee S, Shim JW, Park JH, Lee CJ. Ultrasonic neuromodulation via astrocytic TRPA1. Curr Biol, 29(20), 3386–3401.e8 (2019).
21) Earley S, Gonzales AL, Crnich R. Endothelium-dependent cerebral artery dilation mediated by TRPA1 and Ca2+-activated K+ channels. Circ Res, 104(8), 987–994 (2009).
22) Hamilton NB, Kolodziejczyk K, Kougioumtzidou E, Attwell D. Proton-gated Ca2+-permeable TRP channels damage myelin in conditions mimicking ischaemia. Nature, 529(7587), 523–527 (2016).
23) Pires PW, Earley S. Neuroprotective effects of TRPA1 channels in the cerebral endothelium following ischemic stroke. eLife, 7, e35316 (2018).
24) Lee KI, Lee HT, Lin HC, Tsay HJ, Tsai FC, Shyue SK, Lee TS. Role of transient receptor potential ankyrin 1 channels in Alzheimer’s disease. J Neuroinflammation, 13(1), 92 (2016).
25) Paumier A, Boisseau S, Jacquier-Sarlin M, Pernet-Gallay K, Buisson A, Albrieux M. Astrocyte-neuron interplay is critical for Alzheimer’s disease pathogenesis and is rescued by TRPA1 channel blockade. Brain, 145(1), 388–405 (2022).
26) Ishibashi T, Dakin KA, Stevens B, Lee PR, Kozlov SV, Stewart CL, Fields RD. Astrocytes promote myelination in response to electrical impulses. Neuron, 49(6), 823–832 (2006).
27) Gorelick PB, Scuteri A, Black SE, Decarli C, Greenberg SM, Iadecola C, Launer LJ, Laurent S, Lopez OL, Nyenhuis D, Petersen RC, Schneider JA, Tzourio C, Arnett DK, Bennett DA, Chui HC, Higashida RT, Lindquist R, Nilsson PM, Roman GC, Sellke FW, Seshadri S; American Heart Association Stroke Council, Council on Epidemiology and Prevention, Council on Cardiovascular Nursing, Council on Cardiovascular Radiology and Intervention, and Council on Cardiovascular Surgery and Anesthesia. Vascular contributions to cognitive impairment and dementia: a statement for healthcare professionals from the american heart association/american stroke association. Stroke, 42(9), 2672–2713 (2011).

This page was created on 2024-10-16T16:57:41.059+09:00
This page was last modified on 2024-12-27T08:10:33.000+09:00
このサイトは(株)国際文献社によって運用されています。