グリア由来グルタミン酸トランスポーターのシナプス局在メカニズムの解明
理化学研究所 脳神経科学研究センター グリア-神経回路動態研究チーム
発行日:2024年12月30日Published: December 30, 2024
© 2024 日本神経化学会© 2024 The Japanese Society for Neurochemistry
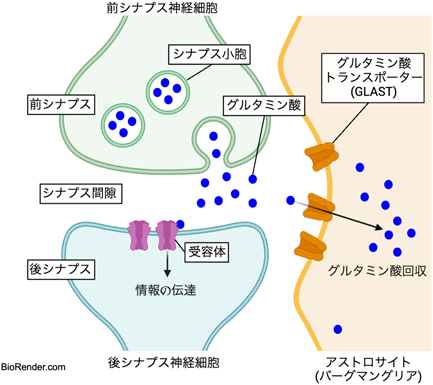
脳神経系は電気的信号により情報を交換する神経細胞のネットワークで形成されている。神経細胞はシナプスと呼ばれる場を介してシナプス前終末から後終末へと、情報の伝達を行う。この際に前終末から分泌される神経伝達物質であるグルタミン酸は、直ちにシナプス周囲を被覆する被覆するグリア細胞(アストロサイト)によって回収される。この除去システムはシナプス機能の恒常性を維持するのに必須であり、遊離グルタミン酸がシナプス間に滞留してしまうと必要以上の神経興奮が起こり、てんかん等の様々な神経疾患を誘発するとされている。
グルタミン酸を細胞内に取り込むシステムとして、細胞膜上に存在するグルタミン酸トランスポーターが存在する(図1)。このトランスポーターは神経細胞とアストロサイトの両方が持っているが、それぞれ役割が異なることが報告されている1)。特にアストロサイト膜上のトランスポーターであるGLASTはより高濃度のグルタミン酸に対して親和性が高く、分泌直後のグルタミン酸を素早く回収する役割を担っている。そしてこの時の回収効率にはGLASTとシナプスの距離が大きく関わっていることが明らかとなっている2)。しかしながらシナプスという非常に微細な構造の中で、GLASTが効率的にシナプスを探し出し、そこに局在するメカニズムについては不明であった。
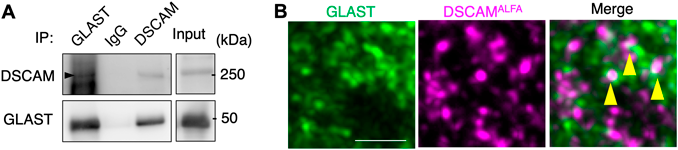
ヒト21番染色体上に存在するDown syndrome cell adhesion molecule (DSCAM)はダウン症の関連遺伝子として発見され3)、近年では統合失調症や自閉症等に関わることも示唆されている4–9)。DSCAMはDSCAM同士の細胞外ドメインを介して接着と反発という異なる機能を使い分けており、これまでさまざまな生物種において神経系の発生・発達に重要な役割を果たしてきていることが報告されてきた10, 11)。また、DSCAMと結合する他の分子も報告されており12)、我々はこの分子の持つ機能的多様性に注目し、小脳シナプス画分からDSCAM抗体を用いた免疫沈降によってDSCAMタンパク質とそれに結合する分子群をまとめて分離し、その分離産物をウエスタンブロット法で調べて見ると、その中にGLAST分子も含まれていることが観察された(図2A)。つまり、小脳においてDSCAMとGLASTが結合していることが示唆された。さらにDSCAM-ALFA tagタンパク質が発現するゲノム編集マウス(DscamALFA/ALFA)の免疫組織化学染色を行った。ALFAタグとGLASTのシグナルが隣接していることから、DSCAM-ALFAとGLASTのシグナルが小脳組織で共局在することが認められた(図2:黄色矢頭)。また、Dscam遺伝子のmRNAの分布を調べたところ、小脳の各種神経細胞では発現するものの、小脳のアストロサイトであるバーグマングリアには発現が認められなかった。そのため、DSCAMとGLASTの相互作用は神経細胞とバーグマングリアの二つの細胞間で起きていると考えられた。
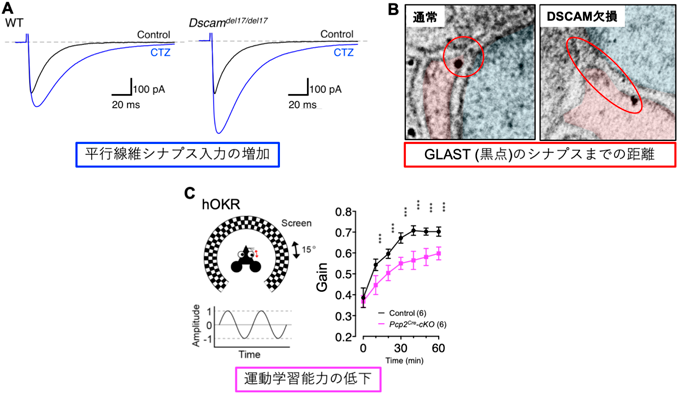
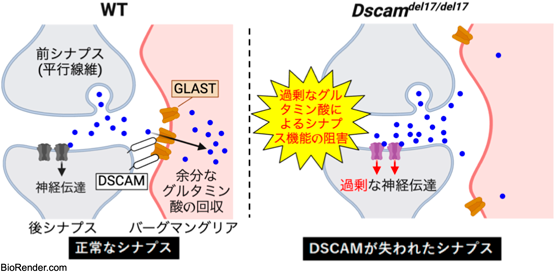
実際にこの相互作用が生体内で機能していることを確かめるため、Dscam遺伝子の機能欠損マウス(Dscamdel17/del17)を用いて以下の実験を行った。まず、成体小脳のプルキンエ細胞に電極を刺してパッチクランプ法で神経活動を調べたところ、Dscamdel17/del17では興奮性シナプスの一つである平行線維シナプスにおいて、GLASTが担っている早い段階でのグルタミン酸回収効率が低下していた(図3A)。さらにこのマウスでは、GLASTタンパク質の発現量自体には変化は認められなかったが、免疫電子顕微鏡実験によってGLASTの興奮性シナプス(平行線維シナプス)における分布を調べると、バーグマングリア細胞膜上のGLAST分子のシナプスへの集積が阻害されていることが明らかになった(図3B)。このことから、神経細胞で発現するDSCAMが失われるとGLASTのシナプス周囲への集積が阻害され、結果的にバーグマングリアによる遊離グルタミン酸の回収が損なわれることが示唆された。
この遊離グルタミン酸除去システムの役割は、過興奮から神経細胞を守ることだけではない。神経発達や学習は、適切なタイミングと適切な閾値での神経伝達によって制御されている。そこで、プルキンエ細胞へと投射する2種類の興奮性シナプス(平行線維シナプスと登上線維シナプス)の発達について調べた。この2種類のシナプスは、お互いに競合してテリトリーを奪い合いながら発達する13)。登上線維シナプスは発達に伴って少しずつプルキンエ細胞の基部から樹状突起の末端方向(上方)へと数を増やしていく一方で、平行線維シナプスはそこから枝分かれした微細な突起でシナプス数を増やすことが知られている13)。正常なマウスと比べて、Dscamdel17/del17マウスでは、登上線維シナプスの上方への拡大が極端に損なわれることが観察された。この現象は、Dscam遺伝子をプルキンエ細胞だけで阻害したノックアウトマウス(Dscamflox/flox;Pfcp2Cre)でも認められたため、プルキンエ細胞で発現するDSCAMタンパク質こそが、このシナプス発達に重要な役割を果たしていることが明らかとなった。この2種類のシナプスが互いのテリトリーを決めるメカニズムには発達時期における平行繊維からの入力が鍵であることが示唆されている14)。DSCAMが失われると平行線維シナプスで遊離グルタミン酸が滞留することによりこのバランスが崩れるため、競合する登上線維シナプスの発達を阻害したという可能性が浮上した。そこで、Dscamflox/flox;Pfcp2Creマウスに対して、GLASTのグルタミン酸取り込み促進剤(リルゾール)を投与したところ、登上線維シナプスの発達異常が軽減された。
最後に、マウスの運動学習(hOKR)について検討した。チェック模様の壁の内側にマウスを固定し、その壁を15度ずつ左右に周期的に動かすと、マウスは目でこの動きを追う。訓練を重ねて学習すると次第にこの動きが上手になってくるが、Dscamflox/flox;Pfcp2Creマウスでは、この学習能力が極端に低下していた(図3C)。登上線維シナプスの発達がこの運動学習に関わることが知られていることから15)、DSCAMが登上線維シナプスの発達制御を介して、運動学習に関与すると考えられる。以上のことから、DSCAM欠損はGLASTによる遊離グルタミン酸の取り込み障害を介した登上線維シナプスの発達異や運動障害を引き起こすことが示唆された(図4)。
グリア由来グルタミン酸トランスポーターにはGLASTとGLT-1の2種類が存在する。アストロサイトがどちらを発現するかは脳領域または脳の状態によって異なる。興味深いことに大脳皮質の抽出液を用いたDSCAMのプロテオミクス結果から、DSCAMはどちらのグルタミン酸トランスポーターとも結合する可能性が示唆された16)。よって今回我々が発見したメカニズムは小脳にとどまらず、より広範囲で機能していると考えられ、他の高次脳機能との関与を明らかにすることで上述の精神疾患の病態の理解に繋がることが期待される。
本研究は、主に国立精神・神経医療研究センター 神経研究所 病態生化学研究部において実施いたしました。特に、当時ご指導を賜りました星野幹雄先生(病態生化学研究部 部長)および、小泉修一先生(山梨大学)には、心より感謝申し上げます。また、論文投稿に至るまで多くの方々から多大なお力添えを賜りましたこと、この場をお借りして厚く御礼申し上げます。さらに、本稿執筆の機会を頂戴いたしました日本神経化学会、ならびに優秀賞・奨励賞選考委員会および編集委員の先生方に、深く感謝申し上げます。今後も、研究者として科学に貢献するべく、また神経化学分野のさらなる発展に寄与するため、一層の努力を重ねる所存です。
1) Takayasu Y, Iino M, Shimamoto K, Tanaka K, Ozawa S. Glial glutamate transporters maintain one-to-one relationship at the climbing fiber-Purkinje cell synapse by preventing glutamate spillover. J Neurosci, 26(24), 6563–6572 (2006).
2) Ageta-Ishihara N, Yamazaki M, Konno K, Nakayama H, Abe M, Hashimoto K, Nishioka T, Kaibuchi K, Hattori S, Miyakawa T, Tanaka K, Huda F, Hirai H, Hashimoto K, Watanabe M, Sakimura K, Kinoshita M. A CDC42EP4/septin-based perisynaptic glial scaffold facilitates glutamate clearance. Nat Commun, 6(1), 10090 (2015).
3) Yamakawa K, Huot YK, Haendelt MA, Hubert R, Chen XN, Lyons GE, Korenberg JR. DSCAM: A novel member of the immunoglobulin superfamily maps in a Down syndrome region and is involved in the development of the nervous system. Hum Mol Genet, 7(2), 227–237 (1998).
4) Amano K, Yamada K, Iwayama Y, Detera-Wadleigh SD, Hattori E, Toyota T, Tokunaga K, Yoshikawa T, Yamakawa K. Association study between the Down syndrome cell adhesion molecule (DSCAM) gene and bipolar disorder. Psychiatr Genet, 18(1), 1–10 (2008).
5) De Rubeis S, He X, Goldberg AP, Poultney CS, Samocha K, Cicek AE, Kou Y, Liu L, Fromer M, Walker S, Singh T, Klei L, Kosmicki J, Shih-Chen F, Aleksic B, Biscaldi M, Bolton PF, Brownfeld JM, Cai J, Campbell NG, Carracedo A, Chahrour MH, Chiocchetti AG, Coon H, Crawford EL, Curran SR, Dawson G, Duketis E, Fernandez BA, Gallagher L, Geller E, Guter SJ, Hill RS, Ionita-Laza J, Jimenz Gonzalez P, Kilpinen H, Klauck SM, Kolevzon A, Lee I, Lei I, Lei J, Lehtimäki T, Lin CF, Ma’ayan A, Marshall CR, McInnes AL, Neale B, Owen MJ, Ozaki N, Parellada M, Parr JR, Purcell S, Puura K, Rajagopalan D, Rehnström K, Reichenberg A, Sabo A, Sachse M, Sanders SJ, Schafer C, Schulte-Rüther M, Skuse D, Stevens C, Szatmari P, Tammimies K, Valladares O, Voran A, Li-San W, Weiss LA, Willsey AJ, Yu TW, Yuen RK, Cook EH, Freitag CM, Gill M, Hultman CM, Lehner T, Palotie A, Schellenberg GD, Sklar P, State MW, Sutcliffe JS, Walsh CA, Scherer SW, Zwick ME, Barett JC, Cutler DJ, Roeder K, Devlin B, Daly MJ, Buxbaum JDDDD StudyHomozygosity Mapping Collaborative for AutismUK10K Consortium. Synaptic, transcriptional and chromatin genes disrupted in autism. Nature, 515(7526), 209–215 (2014).
6) Iossifov I, O’Roak BJ, Sanders SJ, Ronemus M, Krumm N, Levy D, Stessman HA, Witherspoon KT, Vives L, Patterson KE, Smith JD, Paeper B, Nickerson DA, Dea J, Dong S, Gonzalez LE, Mandell JD, Mane SM, Murtha MT, Sullivan CA, Walker MF, Waqar Z, Wei L, Willsey AJ, Yamrom B, Lee YH, Grabowska E, Dalkic E, Wang Z, Marks S, Andrews P, Leotta A, Kendall J, Hakker I, Rosenbaum J, Ma B, Rodgers L, Troge J, Narzisi G, Yoon S, Schatz MC, Ye K, McCombie WR, Shendure J, Eichler EE, State MW, Wigler M. The contribution of de novo coding mutations to autism spectrum disorder. Nature, 515(7526), 216–221 (2014).
7) Yuen CRK, et al. Whole genome sequencing resource identifies 18 new candidate genes for autism spectrum disorder. Nat Neurosci, 20(4), 602–611 (2017).
8) Turner TN, Hormozdiari F, Duyzend MH, McClymont SA, Hook PW, Iossifov I, Raja A, Baker C, Hoekzema K, Stessman HA, Zody MC, Nelson BJ, Huddleston J, Sandstrom R, Smith JD, Hanna D, Swanson JM, Faustman EM, Bamshad MJ, Stamatoyannopoulos J, Nickerson DA, McCallion AS, Darnell R, Eichler EE. Genome sequencing of autism-affected families reveals disruption of putative noncoding regulatory DNA. Am J Hum Genet, 98(1), 58–74 (2016).
9) Wang T, Guo H, Xiong B, Stessman HA, Wu H, Coe BP, Turner TN, Liu Y, Zhao W, Hoekzema K, Vives L, Xia L, Tang M, Ou J, Chen B, Shen Y, Xun G, Long M, Lin J, Kronenberg ZN, Peng Y, Bai T, Li H, Ke X, Hu Z, Zhao J, Zou X, Xia K, Eichler EE. De novo genic mutations among a Chinese autism spectrum disorder cohort. Nat Commun, 7(1), 13316 (2016).
10) Yamagata M, Sanes JR. Dscam and Sidekick proteins direct lamina-specific synaptic connections in vertebrate retina. Nature, 451(7177), 465–469 (2008).
11) Fuerst PG, Koizumi A, Masland RH, Burgess RW. Neurite arborization and mosaic spacing in the mouse retina require DSCAM. Nature, 451(7177), 470–474 (2008).
12) Ly A, Nikolaev A, Suresh G, Zheng Y, Tessier-Lavigne M, Stein E. DSCAM is a netrin receptor that collaborates with DCC in mediating turning responses to netrin-1. Cell, 133(7), 1241–1254 (2008).
13) Kano M, Watanabe T. Developmental synapse remodeling in the cerebellum and visual thalamus. F1000 Res, 8, F1000 (2019).
14) Ichikawa R, Hashimoto K, Miyazaki T, Uchigashima M, Yamasaki M, Aiba A, Kano M, Watanabe M. Territories of heterologous inputs onto Purkinje cell dendrites are segregated by mGluR1-dependent parallel fibre synapse elimination. Proc Natl Acad Sci USA, 113(8), 2282–2287 (2016).
15) Kakegawa W, Mitakidis N, Miura E, Abe M, Matsuda K, Takeo YH, Kohda K, Motohashi J, Takahashi A, Nagao S, Muramatsu S, Watanabe M, Sakimura K, Aricescu AR, Yuzaki M. Anterograde C1ql1 signaling is required in order to determine and maintain a single-winner climbing fiber in the mouse cerebellum. Neuron, 85(2), 316–329 (2015).
16) Arimura N, Okada M, Taya S, Dewa KI, Tsuzuki A, Uetake H, Miyashita S, Hashizume K, Shimaoka K, Egusa S, Nishioka T, Yanagawa Y, Yamakawa K, Inoue YU, Inoue T, Kaibuchi K, Hoshino M. DSCAM regulates delamination of neurons in the developing midbrain. Sci Adv, 6(36), eaba1693 (2020).

This page was created on 2024-10-18T15:58:42.567+09:00
This page was last modified on 2024-12-27T08:18:56.000+09:00
このサイトは(株)国際文献社によって運用されています。