脊髄アストロサイト集団が担う新たな疼痛制御メカニズム
九州大学大学院 薬学研究院 薬理学分野
発行日:2025年6月30日Published: June 30, 2025
© 2025 日本神経化学会© 2025 The Japanese Society for Neurochemistry
グリア細胞の一種であるアストロサイトは、脳や脊髄に一様に存在しており、線虫からヒトに至るまで種々の生物でその存在が確認されている。同細胞は、神経細胞の栄養支持的な役割に加え、近年の研究から細胞外における神経伝達物質の取り込み、またATPやグルタミン酸などのグリア伝達物質の放出を介して神経活動を調節し、行動変容にまで寄与し得ることが明らかとなってきた1)。古典的には、線維状に広がった形態を有し白質に存在する線維性アストロサイト、および星状構造をとり灰白質に存在する原形質アストロサイトに大分され、それぞれ一様な集団であると認識されていた。しかし、近年のシングルセルRNA-seq解析、形態解析およびin vivoのイメージング技術の発達に伴い、アストロサイトの分子発現、形態および細胞応答性は同一領域内あるいは領域間において非常に多様であることが明らかとなっている2)。本稿では、末梢からの感覚情報を受容し、上位中枢に伝達する場として知られる脊髄に存在するアストロサイトの役割について、最新の知見を交えて概説する。
末梢からの感覚情報は、まず始めに一次求心性神経が受容し、脊髄後角へと伝達される。その後、脊髄内の神経回路を介して情報処理を受けた後、上位中枢へと伝達され、我々は感覚を適切に認知すると考えられている3)。
脊髄には神経細胞に加え、グリア細胞が多数存在しており、これまで特に慢性疼痛を含む病態時における役割解析が進んできた。1991年に末梢神経損傷モデルラットの損傷側の脊髄において、グリア線維性酸性タンパク質(glial fibrillary acidic protein: GFAP)の発現増加が観察されたことにより、神経障害性疼痛モデルにおいて脊髄アストロサイトの活性化が初めて示された4)。その後、神経損傷や炎症性疼痛モデルを含む様々な慢性疼痛モデルにおいて、グリア細胞に選択性のある代謝阻害剤(fluorocitrate5))やアストロサイト毒素(L-α-aminoadipate6))などを用いて活性化を抑制した際に、アロディニア(通常は痛みを感じない軽微な刺激を痛みとして感じる感覚異常)の改善が観察され、慢性疼痛病態への関与が示唆されることになる。
さらに、脊髄アストロサイトの病態への関与を詳細に検討するため、同細胞で特異的に活性化する細胞内シグナルを同定する研究が進む。中でもMitogen-activated protein kinase(MAPK)の一種であるERK7)やJNK6)の活性化が慢性疼痛病態モデルにおける脊髄アストロサイトで観察され、それらに対する阻害剤の投与により慢性疼痛が緩和したことから、疾患の治療標的としてアストロサイトに注目が集まることになる。そのような中、筆者らは末梢神経損傷モデルラットにおいて、転写因子の1種であるSTAT3が脊髄アストロサイトで活性化し、細胞増殖を介して慢性疼痛に重要であることを報告した8)。また、神経障害性疼痛に加え、脊柱管狭窄症に代表される脊髄圧迫時の疼痛病態にも脊髄アストロサイトのSTAT3シグナルが重要であることを明らかとしている9)。その後の研究により、種々のMAPKや転写因子を介して発現増加したサイトカイン(TNF-α, IL-1βなど)やケモカイン(CCL2, CXCL1など)が脊髄神経細胞に作用し、神経活動を変調させることで、慢性疼痛病態へと関与すると考えられている10)。
病態時における脊髄アストロサイトの役割解明が進む一方で、正常時の感覚情報伝達における同細胞の存在意義については不明なままであった。そのような中、開口放出に重要なSNARE複合体のドミナントネガティブ体をアストロサイト特異的に発現させたマウスが痛覚過敏を示すことが2011年に報告され、病態時のみならず正常時においても、脊髄アストロサイトが末梢からの感覚情報伝達の処理に関与する可能性が示された11)。2016年には、光活性型イオンチャネルの一種であるChR2を脊髄アストロサイトに発現させたラットを用いて、腰髄のアストロサイトを選択的に光刺激することで痛覚過敏が生じることが報告された12)。一方で、マウス足裏への電気刺激によりAβ線維を選択的に刺激した際に脊髄アストロサイトでカルシウム応答が観察され、同反応が鎮痛効果を生じることも示されている13)。一連の報告から、正常時の感覚情報伝達における脊髄アストロサイトの役割は、活動性が変化する集団によりその影響力が異なる可能性が考えられる。近年の研究により、脳アストロサイトは分子発現、形態および機能が多様であることが明らかとなっており、同一領域内においても集団により細胞機能が異なることが示されている。一方で、脊髄アストロサイトの多様性、および各集団がどのように感覚変調に寄与し得るのかは不明であった10)。下記では、筆者らがこれまで明らかにしてきた、正常時の感覚情報伝達に対する脊髄アストロサイト集団の役割およびそのメカニズムについて詳述する。
筆者らは、解剖学的に脊髄後角と前角を分け、既報のアストロサイト関連分子について遺伝子発現解析を行ったところ、転写因子の一種であるhairy and enhancer of split 5(Hes5)が脊髄後角において優位に発現していることを見出した。また、Hes5陽性細胞で蛍光タンパク質のtdTomatoを発現する遺伝子改変マウスを用いることで、脊髄後角表層(Lamina I-IIIo)に存在するアストロサイトがHes5陽性であることを同定した。同細胞が局在する領域は、末梢からの侵害刺激が入力する場所であることから、マウス足裏へカプサイシンを投与した際の同細胞におけるカルシウム応答についてin vivo脊髄イメージング14)を用いて解析したところ、細胞内カルシウム濃度増加が観察された。また、興味深いことに、脊髄の観察部位とは反対側の足裏にカプサイシンを投与した際もカルシウム応答が惹起された。同結果から、一次求心性神経のみならず他領域からのシグナルがアストロサイトのカルシウム応答を惹起する可能性が考えられたため、脳からの下行性疼痛制御系として代表的なセロトニンおよびノルアドレナリンシグナルの関与について検討を行った。急性単離脊髄スライスを用いてアストロサイト細胞内カルシウム応答について検討を行ったところ、セロトニン処置時は応答がなかったのに対し、ノルアドレナリン処置によりカルシウム応答が惹起された。各種阻害剤を用いて責任受容体の探索を行い、Hes5陽性アストロサイトに発現するα1A受容体がノルアドレナリン誘発のカルシウム応答に重要であることを明らかにした。
そこで次に、アストロサイトの細胞内カルシウム濃度増加が感覚情報伝達に及ぼす影響について検討した。アデノ随伴ウイルス(AAV)を用いて、人工受容体として知られるhM3Dq15)を脊髄Hes5陽性アストロサイト特異的に発現させたマウスを作製し16)、選択的な刺激を行ったところ、機械刺激に対する過敏応答を惹起した。さらに同過敏応答は、NMDA受容体のグリシン結合部位に対する阻害剤の前処置により抑制され、また同部位へ結合可能なD-セリンを脊髄腔内に投与することで痛覚過敏が生じた。一方で、脊髄Hes5陰性アストロサイトを選択的に刺激した際は、感覚過敏は惹起されなかった。以上の結果より、Hes5陽性アストロサイト内でのカルシウム濃度増加によりD-セリンが放出され、痛覚過敏を惹起する可能性が示された。
マウス足裏にカプサイシンを投与すると、即時的な侵害防御行動(肢をなめる、噛むなど)に加え、機械刺激に対する過敏応答が遅延して生じる。そこで、Hes5陽性アストロサイト特異的α1A受容体ノックアウトマウスを作製し、カプサイシン投与後の行動解析を行ったところ、野生型マウスと比較して侵害防御行動は同程度惹起されたのに対し、痛覚過敏が抑制された。さらに、青斑核のノルアドレナリン神経を選択的に変性可能なDSP-417)を処置したマウスでも、カプサイシン誘発のカルシウム応答および痛覚過敏が抑制された18)。
以上の結果から、これまで下行性疼痛抑制系として考えられてきた青斑核由来のノルアドレナリンシグナルは、脊髄アストロサイト集団を介することで疼痛促進系としても機能することを明らかにした。侵害刺激受容後の過敏応答は、神経細胞ではなし得ない一種の生体防御機構であると考えられ、感覚情報伝達における脊髄アストロサイト集団の存在意義の一端を明らかにしたと言える。
青斑核は種々の外的刺激に反応し、個体の行動を変容する。急性ストレス刺激は青斑核を活性化させること19)、またストレスが慢性疼痛の憎悪にも関与すること20)が示唆されているがその詳細なメカニズムは不明である。そこで、急性ストレス負荷時の行動変容について、筆者らが同定した下行性ノルアドレナリン神経–脊髄Hes5陽性アストロサイトの関与について検討を行った。マウスに拘束ストレスを1時間負荷すると痛覚過敏が生じ、この時、ファイバーフォトメトリーを用いて青斑核の神経活動が亢進していることが確認された。また、AAVを用いて脊髄に投射する青斑核のノルアドレナリン神経を変性させると拘束ストレス誘発の過敏応答が減弱した。一方で、光刺激により同神経を活性化することで、痛覚過敏が惹起された。さらに、Hes5陽性アストロサイト特異的α1A受容体のノックアウトマウスでは痛覚過敏が消失した。そこで、ストレス負荷時の脊髄での応答について観察するため、脊髄へファイバーフォトメトリーを留置し、拘束ストレス下での反応について解析したところ、脊髄でのノルアドレナリン放出およびアストロサイトのカルシウム応答が観察された。以上の結果より、拘束ストレスにより青斑核ノルアドレナリン神経–脊髄Hes5陽性アストロサイトを介して痛覚過敏が惹起されることが明らかとなった。
筆者の所属研究室は、脊髄アストロサイトと同様に、抑制性の介在神経にもα1A受容体が発現することを報告している21)。しかしながら、抑制性神経特異的α1A受容体ノックアウトマウスでは、拘束ストレス誘発の痛覚過敏に影響がなかった。そこで次に、Hes5陽性アストロサイト活性化時に抑制性神経の活動性が変化するのではないかという仮説を立て、電気生理学的な解析を行った。過去に、脊髄アストロサイトをChR2により光刺激することでATPが放出され、分解されたアデノシンが抑制性神経に作用することが報告されており12)、ノルアドレナリン処置による抑制性神経の活動性亢進に対するアデノシンシグナルの影響について検討した。その結果、ノルアドレナリン誘発の抑制性神経の脱分極応答は、アデノシンA1Rのアゴニスト処置により抑制された。また、SaCas9を搭載したAAVベクターによるゲノム編集を利用して、抑制性神経特異的にA1Rをノックアウトしたマウスを作製したところ、アデノシンA1Rのアゴニストによる抑制効果が消失した。さらに、Hes5陽性アストロサイトをhM3Dqで選択的に刺激すると、膠様質の神経細胞におけるsIPSCが減弱し、この効果はA1Rの阻害剤処置により抑制された。上述の結果を踏まえ、ストレス誘発性痛覚過敏におけるアデノシンシグナルの関与について検討を行ったところ、A1Rの阻害剤の前処置、あるいは抑制性神経特異的なA1Rノックアウトにより拘束ストレス誘発の痛覚過敏が消失した22)。
以上の結果から、脊髄Hes5陽性アストロサイトはストレスを痛みへと変換することで、生体における警告系として重要な役割を果たしていると考えられる。また、青斑核由来のノルアドレナリンによる脊髄アストロサイト–抑制性神経の相互作用を介した痛覚過敏は、痛覚変調性疼痛をはじめとした慢性疼痛の治療標的となることが期待される(図1)。
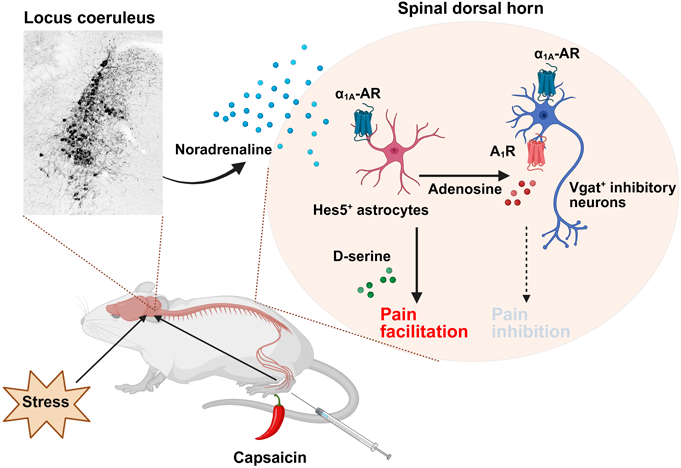
末梢への侵害刺激(足裏へのカプサイシン投与)あるいは急性ストレス刺激により、青斑核(Locus coeruleus)のノルアドレナリン神経が活性化する。脊髄後角(spinal dorsal horn)においてノルアドレナリンが放出され、Hes5陽性アストロサイトのα1A受容体に作用し、D-セリンを介して疼痛促進へと働く。この時、Hes5陽性アストロサイトにより抑制性神経のアデノシンシグナルが活性化し、同神経の神経活動が抑制される。BioRender.comを用いて図を作製。
GFAPの発現変化を軸としたアストロサイト研究は、イメージング技術の発達や、シングルセルRNA-seq解析などの技術進歩に伴い、様々な角度から解析が可能となり、その細胞機能の多様性が明らかとなってきている。本稿では、脊髄Hes5陽性アストロサイト集団について詳述したが、最近では脊髄後角深層に存在するLfng-GFP陽性アストロサイト(Lamina III–IV)が特に軽微な機械刺激に対する応答に重要であることも報告され23)、脊髄アストロサイトの機能的な多様性が示唆されている。また、筆者らは脊髄アストロサイトの選択的かつ効率的な単離法を確立しており24)、シングルセルRNA-seq解析を用いてさらなる特徴的な集団の同定および機能解析を進めている。今後、新たな脊髄アストロサイト集団を介する感覚情報伝達制御メカニズムや、病態時における脊髄アストロサイト集団の役割が解明され、慢性疼痛を含む疾患の治療標的に資する細胞集団や分子の同定が期待される。
本研究を行うにあたり、多大なるご指導とご協力を賜りました九州大学高等研究院 井上和秀 特別主幹教授、九州大学大学院薬学研究院 津田誠 主幹教授、川邉陸 博士、内山瑳和子 修士、吉原康平 博士、および研究室の皆様、共同研究者の先生方にこの場を借りて深く感謝申し上げます。また、この度の執筆の機会を与えていただきました日本神経化学会出版・広報委員会の澤本和延委員長、委員の先生ならびに編集部の皆様に深く御礼申し上げます。
1) Nagai J, Yu X, Papouin T, Cheong E, Freeman MR, Monk KR, Hastings MH, Haydon PG, Rowitch D, Shaham S, Khakh BS. Behaviorally consequential astrocytic regulation of neural circuits. Neuron, 109(4), 576–596 (2021).
2) Ben Haim L, Rowitch DH. Functional diversity of astrocytes in neural circuit regulation. Nat Rev Neurosci, 18(1), 31–41 (2017).
3) Peirs C, Seal RP. Neural circuits for pain: Recent advances and current views. Science, 354(6312), 578–584 (2016).
4) Garrison CJ, Dougherty PM, Kajander KC, Carlton SM. Staining of glial fibrillary acidic protein (GFAP) in lumbar spinal cord increases following a sciatic nerve constriction injury. Brain Res, 565(1), 1–7 (1991).
5) Meller ST, Dykstra C, Grzybycki D, Murphy S, Gebhart GF. The possible role of glia in nociceptive processing and hyperalgesia in the spinal cord of the rat. Neuropharmacology, 33(11), 1471–1478 (1994).
6) Zhuang ZY, Wen YR, Zhang DR, Borsello T, Bonny C, Strichartz GR, Decosterd I, Ji RR. A peptide c-Jun N-terminal kinase (JNK) inhibitor blocks mechanical allodynia after spinal nerve ligation: respective roles of JNK activation in primary sensory neurons and spinal astrocytes for neuropathic pain development and maintenance. J Neurosci, 26(13), 3551–3560 (2006).
7) Zhuang ZY, Gerner P, Woolf CJ, Ji RR. ERK is sequentially activated in neurons, microglia, and astrocytes by spinal nerve ligation and contributes to mechanical allodynia in this neuropathic pain model. Pain, 114(1–2), 149–159 (2005).
8) Tsuda M, Kohro Y, Yano T, Tsujikawa T, Kitano J, Tozaki-Saitoh H, Koyanagi S, Ohdo S, Ji RR, Salter MW, Inoue K. JAK-STAT3 pathway regulates spinal astrocyte proliferation and neuropathic pain maintenance in rats. Brain, 134(Pt 4), 1127–1139 (2011).
9) Ono T, Kohro Y, Kohno K, Tozaki-Saitoh H, Nakashima Y, Tsuda M. Mechanical pain of the lower extremity after compression of the upper spinal cord involves signal transducer and activator of transcription 3-dependent reactive astrocytes and interleukin-6. Brain Behav Immun, 89, 389–399 (2020).
10) Ji RR, Donnelly CR, Nedergaard M. Astrocytes in chronic pain and itch. Nat Rev Neurosci, 20(11), 667–685 (2019).
11) Foley JC, McIver SR, Haydon PG. Gliotransmission modulates baseline mechanical nociception. Mol Pain, 7, 93 (2011).
12) Nam Y, Kim JH, Kim JH, Jha MK, Jung JY, Lee MG, Choi IS, Jang IS, Lim DG, Hwang SH, Cho HJ, Suk K. Reversible induction of pain hypersensitivity following optogenetic stimulation of spinal astrocytes. Cell Rep, 17(11), 3049–3061 (2016).
13) Xu Q, Ford NC, He S, Huang Q, Anderson M, Chen Z, Yang F, Crawford LK, Caterina MJ, Guan Y, Dong X. Astrocytes contribute to pain gating in the spinal cord. Sci Adv, 7(45), eabi6287 (2021).
14) Yoshihara K, Matsuda T, Kohro Y, Tozaki-Saitoh H, Inoue K, Tsuda M. Astrocytic Ca2+ responses in the spinal dorsal horn by noxious stimuli to the skin. J Pharmacol Sci, 137(1), 101–104 (2018).
15) Roth BL. DREADDs for neuroscientists. Neuron, 89(4), 683–694 (2016).
16) Kohro Y, Sakaguchi E, Tashima R, Tozaki-Saitoh H, Okano H, Inoue K, Tsuda M. A new minimally-invasive method for microinjection into the mouse spinal dorsal horn. Sci Rep, 5(1), 14306 (2015).
17) Ross SB, Stenfors C. DSP4, a selective neurotoxin for the locus coeruleus noradrenergic system. A review of its mode of action. Neurotox Res, 27(1), 15–30 (2015).
18) Kohro Y, Matsuda T, Yoshihara K, Kohno K, Koga K, Katsuragi R, Oka T, Tashima R, Muneta S, Yamane T, Okada S, Momokino K, Furusho A, Hamase K, Oti T, Sakamoto H, Hayashida K, Kobayashi R, Horii T, Hatada I, Tozaki-Saitoh H, Mikoshiba K, Taylor V, Inoue K, Tsuda M. Spinal astrocytes in superficial laminae gate brainstem descending control of mechanosensory hypersensitivity. Nat Neurosci, 23(11), 1376–1387 (2020).
19) Valentino RJ, Van Bockstaele E. Convergent regulation of locus coeruleus activity as an adaptive response to stress. Eur J Pharmacol, 583(2–3), 194–203 (2008).
20) Fitzcharles MA, Cohen SP, Clauw DJ, Littlejohn G, Usui C, Häuser W. Nociplastic pain: towards an understanding of prevalent pain conditions. Lancet, 397(10289), 2098–2110 (2021).
21) Uchiyama S, Yoshihara K, Kawanabe R, Hatada I, Koga K, Tsuda M. Stress-induced antinociception to noxious heat requires α1A-adrenaline receptors of spinal inhibitory neurons in mice. Mol Brain, 15(1), 6 (2022).
22) Kawanabe-Kobayashi R, Uchiyama S, Yoshihara K, Kojima D, McHugh TJ, Hatada I, Matsui K, Tanaka KF, Tsuda M. Descending locus coeruleus noradrenergic signaling to spinal astrocyte subset is required for stress-induced pain facilitation. eLife, (2025). https://doi.org/10.7554/eLife.104453.1
23) Akdemir ES, Woo J, Bosquez Huerta NA, Lozzi B, Groves AK, Harmanci AS, Deneen B. Lunatic Fringe-GFP marks lamina-specific astrocytes that regulate sensory processing. J Neurosci, 42(4), 567–580 (2022).
24) Iwasaki R, Kohro Y, Tsuda M. A method for selective and efficient isolation of gray matter astrocytes from the spinal cord of adult mice. Mol Brain, 17(1), 25 (2024).

This page was created on 2025-07-10T17:13:46.177+09:00
This page was last modified on 2025-07-25T14:48:22.000+09:00
このサイトは(株)国際文献社によって運用されています。