生後の新生ニューロン移動による脳傷害後の神経再生
1 名古屋大学大学院 理学研究科 附属ニューロサイエンス研究センター 細胞制御学グループ
2 名古屋市立大学大学院 医学研究科 脳神経科学研究所 神経発達・再生医学分野
発行日:2025年6月30日Published: June 30, 2025
© 2025 日本神経化学会© 2025 The Japanese Society for Neurochemistry
脳梗塞などの脳損傷によって神経細胞(ニューロン)やグリア細胞は失われ、脳機能障害が生じる。一方で、生後の脳においても側脳室下帯(ventricular-subventricular zone, V-SVZ)においては新生ニューロンが産生されることから、脳は損傷後に機能回復できるポテンシャルを備えている。
新生ニューロンは脳内の損傷部に向かって移動し、定着した脳領域において成熟する。筆者らはこの内在的なニューロン再生機能を活用することで、損傷後の脳機能の回復が可能であることを示した。新生ニューロンの移動は損傷後の脳領域では阻まれるが、筆者らはニューロン移動の抑制および促進を規定する分子を特定し、新生ニューロン移動を促進するバイオマテリアルを利用することで、脳再生を可能とした。本稿では、これまでに明らかになった傷害脳における新生ニューロン移動の分子機序と開発したバイオマテリアルについて概説する。
ヒトを含む霊長類、齧歯類など多様な動物種の生後の脳では、海馬歯状回の顆粒細胞下層とV-SVZに神経幹細胞が存在し、新生ニューロンを産生する1–3)。V-SVZ由来の新生ニューロンは、主に吻側移動流を通り、嗅球に向かって移動し、嗅覚機能に関与する抑制性介在ニューロンに分化する4)。新生ニューロンは移動する時には、新生ニューロン同士、あるいは周囲のグリア細胞などの足場細胞と接着と乖離を繰り返して鎖状移動しており5)、細胞間相互作用は移動に重要である。目的とする脳領域に向かう新生ニューロンの鎖状移動は魚類から霊長類まで広く保存されており、普遍的な生命現象であると言える6)。
大脳皮質あるいは線条体などに損傷が生じると、V-SVZの神経幹細胞は休止期を離脱し、活性化状態をとり、ニューロン新生が亢進する7)。産生された新生ニューロンは嗅球の他に、血管やグリア細胞を足場にして損傷部位へ向かうことが明らかになっている8)。脳の発生過程において放射状グリア細胞は脳室面側から脳表面である軟膜側に向かって長い突起を伸ばし、極性を持つ。新生ニューロンは胎生期において放射状グリア細胞の突起を足場として軟膜側に向かって移動する9)。哺乳類では本来、放射状グリア細胞は出生直後に上衣細胞や神経幹細胞に分化し消失するが、大脳皮質の脳傷害モデルマウスでは生後であっても放射状グリア細胞は一定期間維持される10)。そして、傷害脳における新生ニューロンは細胞接着分子N-cadherinを発現し、N-cadherinを発現する放射状グリア細胞の突起に沿って傷害を受けた大脳皮質に向かって移動する10)(図1A)。このようにして傷害脳においては正常脳と同様に、内在する細胞間における相互作用が機能し、損傷部位において失われたニューロンを補おうとする自発的な回復能力を備えている。また、新生ニューロンは胎生期と生後の傷害脳において、共通してN-cadherinを発現する放射状グリア細胞を介して目的とする脳領域に向かって移動することから、新生ニューロンの移動機構として極性を持った放射状グリア細胞を足場とすることは、回復において重要であることが考えられる。
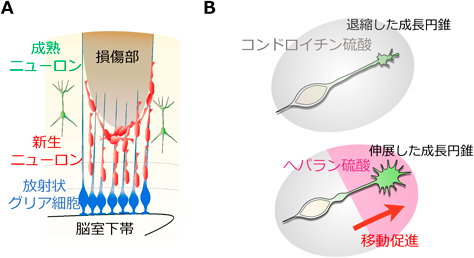
損傷部位では活性化したグリア細胞や血管内皮細胞から炎症性サイトカインやケモカインなどが分泌される。新生ニューロンはこれらの受容体を発現することから、ケモカインなどは誘引に関わることが示唆されている8)。一方で、傷害脳は上述の脳の自発的な回復能力だけでは自然に機能回復しない。傷害脳の機能回復が難しい理由として、十分な量の新生ニューロンが離れた損傷部に移動できないことが考えられる。実際に、新生ニューロンは損傷部位に向かう過程で、移動を停止、あるいはV-SVZに引き返す現象が見られる11–13)。筆者は、大脳皮質傷害モデルマウスの脳切片を用いてスライス培養し、新生ニューロンの突起先端の構造物が伸展および退縮を繰り返しながら損傷部位に向かって移動する様相を捉え、一定距離を移動すると突起先端がV-SVZ方向に反転して、損傷部位から離れて移動することを明らかにした13)。また、脳損傷部近傍においてはV-SVZ近傍と比較して、突起先端の構造物、すなわち成長円錐が退縮することを示した13)。
筆者らは、成長円錐が新生ニューロンの突起先端に存在し、周囲の細胞外マトリックスを感知しながら、細胞骨格を動的に制御することで、移動を調節していることを見出した13)。成長円錐は受容体PTPσを発現し、損傷部位に蓄積する細胞外マトリックスの一つであるコンドロイチン硫酸プロテオグリカン(CSPG)を認識することがわかった。CSPGが成長円錐の退縮を引き起こし、ニューロン移動を抑制しているメカニズムを明らかにした。また、PTPσはへパラン硫酸プロテオグリカン(HSPG)の受容体でもあり、CSPGによる抑制効果をHSPGによって緩和し、成長円錐を伸展ならびに新生ニューロンの移動を促進することが可能となった。成長円錐のフィロポディア突起がニューロン移動の方向性を決定することも示した。以上の結果から、新生ニューロンの成長円錐が、周囲の環境を認識するアンテナであり、ニューロン移動を制御する装置であることが示された13)(図1B)。傷害脳の損傷部において成長円錐の機能を活用することはニューロン移動を制御する上で不可欠である。
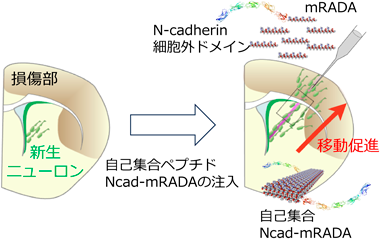
脳損傷の治療法を開発する上で、足場細胞を模倣した人工足場バイオマテリアルを損傷部に移植することが考えられる。これまでに脳表層部の損傷において、新生ニューロンは移植したN-cadherin10)やLaminin14)を含むゼラチンスポンジを移動の足場とすることが明らかになった10)。しかし、スポンジ材料は脳深部の傷害に対しては侵襲性が大きく適応が困難であった。そこで今回、筆者らは注入型ペプチド材料を活用した15)。ペプチド材料mRADAは生体内条件下で自己集合して繊維状のハイドロゲルを形成する。mRADAは脳深部に注入でき、従来のスポンジ材料と比べて侵襲性が少ない他、極性を持った放射状グリア細胞を模してV-SVZから脳表層まで連なった足場を作成することが可能となった。さらに、自己集合性ペプチドmRADAにN-cadherinの細胞外ドメインを組み込むこみ、放射状グリア細胞の形状と発現分子を模倣したNcad-mRADAを開発した。新生仔大脳皮質傷害モデルマウスにおいて新生ニューロンはNcad-mRADAを足場として、脳室下帯から離れた損傷部位まで移動し、再生した成熟ニューロンが損傷領域において増加した(図2)。さらに、新生仔マウスでは歩行機能も回復し、注入型ペプチド材料Ncad-mRADAによる治療効果を示した。また、ハイドロゲルは生体内において分解されることを確認し、再生医療に活用が可能と考えられた。成体脳梗塞モデルマウスにおいてもmRADA注入群と比較してNcad-mRADA注入群は、産生した新生ニューロンが有意に多くかつ遠くまでペプチド材料の足場に沿って移動できた。放射状グリア細胞は本来、出生直後で消失することから、Ncad-mRADAが成体マウス脳において足場として有用であるか不明であった。しかし、今回の結果から成体マウスの新生ニューロンは発生過程や生後直後の新生ニューロンの特性を維持しており、接着分子を組み込んだ足場材料と相互作用することが示された15)。本マテリアルはこれまでニューロンの移動と再生が困難であった脳深部においてもアプローチを可能とし、さまざまな分子を組み込めることから、適用範囲が広く有望なバイオマテリアルである。
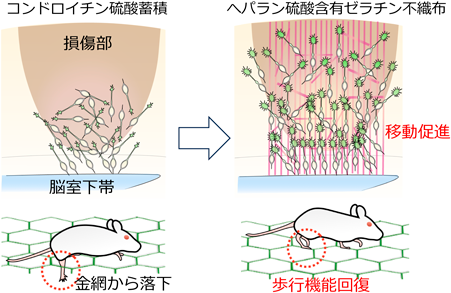
筆者らは続いて、ゼラチン不織布を足場材料として活用した脳機能回復法を開発した13)。ゼラチンはタンパク質との親和性が高く、生体適合性に優れている。用いたゼラチン不織布は網目状にゼラチン繊維が織り込まれた高い連通性と配向性を有する特徴を持つ。ゼラチン繊維間で空間が保持され、複数の繊維が主に一方向に配向することから、傷害脳に移植したゼラチン不織布は新生ニューロンの移動を妨げず、脳表面に到達する放射状グリア細胞の代用となる。
筆者は、新生仔大脳皮質傷害モデルマウスを用いて、CSPGが蓄積する損傷部位に、リコンビナントHSPGを含むゼラチン不織布を移植した。培養条件下において新生ニューロンの移動促進効果を確認できたリコンビナントHSPGの中から、グリピカン4(GPC4)、シンデカン2(SDC2)、およびシンデカン4(SDC4)を選択して、脳傷害モデルマウスの実験に用いた。傷害脳において、新生ニューロンの成長円錐はいずれのHSPGを含むゼラチン繊維に接触すると伸展し、ニューロン移動が促進することをスライス培養から明らかにした。新生ニューロンはHSPG含有ゼラチン不織布を移植すると1週間後には大脳皮質の下層から上層まで広く分布した。移植1ヶ月後の大脳皮質では、HSPG含有群においては再生した成熟ニューロンの数が増加し、さらにマウスの歩行機能も改善した(図3)。遺伝学的手法を用いて、生後に産生した新生ニューロンを除去すると、HSPG含有群で歩行機能の改善効果が消失したことから、新生ニューロンが神経機能改善を担っていたと考えられる。また、HSPG含有ゼラチン不織布の繊維が脳梁に沿うように損傷部に移植して、脳表面に向かって伸びる繊維を少なくすると、大脳皮質の表層部に到達する新生ニューロンの数が減少した。そして、HSPGを付加したポリプロピレン不織布を移植した場合、移動した新生ニューロンは主に第5–6層に観察され、分布が狭い範囲に限定された。以上から、新生ニューロンの移動が抑制されるCSPGが蓄積する環境下においても、適切な人工足場に含ませたHSPGによって新生ニューロンの成長円錐を制御することで、抑制を解除することが可能となることを見出した13)。本研究から、脳損傷後の神経機能改善における新生ニューロンの有用性ならびに、バイオマテリアルの選択の重要性が明らかになった。生後に限らず、胎生期の新生ニューロンの成長円錐もPTPσを発現することから13)、発生初期からの脳形成においてもプロテオグリカンの関与が示唆され、脳形成障害における治療介入法の開発に本研究の有効性が考えられる。
本稿では、生体内における新生ニューロンの移動メカニズムとバイオマテリアルの応用による傷害後の脳機能回復法を概説した。新生ニューロンの移動における接着分子を介した足場細胞との相互作用と、成長円錐の動態を制御する分子メカニズムを理解することで、これまで困難であった傷害脳における新生ニューロンの移動の改善が可能となった。そして、共同研究者とのバイオマテリアルの開発を通じ、脳深部にアプローチできる注入型ペプチド材料と、脳表層の広範囲にわたり足場を供給できるゼラチン不織布が足場材料として効果的であることを示した。新生ニューロンの移動は生体内における周囲の細胞ならびに分泌タンパク質に寄るところが大きいことを本研究から明らかにしたが、新生ニューロン自身も傷害脳においては変化し、鎖状移動に必須の細胞接着が破綻することが近年明らかになった16)。今後も生後の脳機能を司る新生ニューロンの機能解明を通じて、学術的な発展が期待され、再生医療応用にむけて基盤が確立すると考えられる。
近年、ヒトの脳においても神経幹細胞がV-SVZに存在しており、生後1.5年程度までニューロン新生が見られ、新生ニューロンが前頭葉に移動することが明らかになった17)。また、ヒトでも脳梗塞後にV-SVZにおける新生ニューロンの産生が亢進し18)、損傷部に新生ニューロンが確認されている19)。これらの知見から、ヒトの脳にも齧歯類と同様に内在的なニューロン再生機能を有することが示唆される。これまでのニューロン再生研究は主に齧歯類を用いて行われているが、ヒトの脳内における生後のニューロン新生のメカニズムならびに新生ニューロンの機能と意義を理解することで、ヒトの脳の可塑性や再生に通じる研究が活性化すると期待される。
本研究を行うにあたり、多大なるご指導を賜りました名古屋市立大学の澤本和延先生、同志社大学の金子奈穂子先生、研究室の皆様、ならびに多くの共同研究者の先生方に、この場を借りて深く御礼申し上げます。また、この度はこのような執筆の機会を与えてくださいました日本神経化学会出版・広報委員会の諸先生方ならびに編集部の皆様に、深く御礼申し上げます。
1) Doetsch F, Caille I, Lim DA, Garcia-Verdugo JM, Alvarez-Buylla A. Subventricular zone astrocytes are neural stem cells in the adult mammalian brain. Cell, 97(6), 703–716 (1999).
2) Fuentealba LC, Rompani SB, Parraguez JI, Obernier K, Romero R, Cepko CL, Alvarez-Buylla A. Embryonic origin of postnatal neural stem cells. Cell, 161(7), 1644–1655 (2015).
3) Furutachi S, Miya H, Watanabe T, Kawai H, Yamasaki N, Harada Y, Imayoshi I, Nelson M, Nakayama KI, Hirabayashi Y, Gotoh Y. Slowly dividing neural progenitors are an embryonic origin of adult neural stem cells. Nat Neurosci, 18(5), 657–665 (2015).
4) Lois C, Alvarez-Buylla A. Long-distance neuronal migration in the adult mammalian brain. Science, 264(5162), 1145–1148 (1994).
5) Lois C, Garcia-Verdugo JM, Alvarez-Buylla A. Chain migration of neuronal precursors. Science, 271(5251), 978–981 (1996).
6) Sawada M, Sawamoto K. Mechanisms of neurogenesis in the normal and injured adult brain. Keio J Med, 62(1), 13–28 (2013).
7) Chavali M, Klingener M, Kokkosis AG, Garkun Y, Felong S, Maffei A, Aguirre A. Non-canonical Wnt signaling regulates neural stem cell quiescence during homeostasis and after demyelination. Nat Commun, 9(1), 36 (2018).
8) Nakajima C, Sawada M, Sawamoto K. Postnatal neuronal migration in health and disease. Curr Opin Neurobiol, 66, 1–9 (2021).
9) Rakic P. Mode of cell migration to the superficial layers of fetal monkey neocortex. J Comp Neurol, 145(1), 61–83 (1972).
10) Jinnou H, Sawada M, Kawase K, Kaneko N, Herranz-Pérez V, Miyamoto T, Kawaue T, Miyata T, Tabata Y, Akaike T, Garcia-Verdugo JM, Ajioka I, Saitoh S, Sawamoto K. Radial glial fibers support neuronal migration and regeneration after neonatal brain injury. Cell Stem Cell, 22, 128–137 (2018).
11) Kojima T, Hirota Y, Ema M, Takahashi S, Miyoshi I, Okano H, Sawamoto K. Subventricular zone-derived neural progenitor cells migrate along a blood vessel scaffold toward the post-stroke striatum. Stem Cells, 28(3), 545–554 (2010).
12) Kaneko N, Herranz-Pérez V, Otsuka T, Sano H, Ohno N, Omata T, Nguyen HB, Thai TQ, Nambu A, Kawaguchi Y, García-Verdugo JM, Sawamoto K. New neurons use Slit-Robo signaling to migrate through the glial meshwork and approach a lesion for functional regeneration. Sci Adv, 4(12), eaav0618 (2018).
13) Nakajima C, Sawada M, Umeda E, Takagi Y, Nakashima N, Kuboyama K, Kaneko N, Yamamoto S, Nakamura H, Shimada N, Nakamura K, Matsuno K, Uesugi S, Veprek N, Küllmer F, Nasufovic V, Uchiyama H, Nakada M, Otsuka Y, Ito Y, Herranz-Pérez V, García-Verdugo JM, Ohno N, Arndt HD, Trauner D, Tabata Y, Igarashi M, Sawamoto K. Identification of the growth cone as a probe and driver of neuronal migration in the injured brain. Nat Commun, 15(1), 1877 (2024).
14) Ajioka I, Jinnou H, Okada K, Sawada M, Saitoh S, Sawamoto K. Enhancement of neuroblast migration into the injured cerebral cortex using laminin-containing porous sponge. Tissue Eng Part A, 21(1–2), 193–201 (2015).
15) Ohno Y, Nakajima C, Ajioka I, Muraoka T, Yaguchi A, Fujioka T, Akimoto S, Matsu M, Lotfy A, Nakamura S, Herranz-Pérez V, García-Verdugo JM, Matsukawa N, Kaneko N, Sawamoto K. Amphiphilic peptide-tagged N-cadherin forms radial glial-like fibers that enhance neuronal migration in injured brain and promote sensorimotor recovery. Biomaterials, 294, 122003 (2023).
16) Matsumoto M, Matsushita K, Hane M, Wen C, Kurematsu C, Ota H, Nguyen HB, Thai TQ, Herranz-Pérez V, Sawada M, Fujimoto K, García-Verdugo JM, Kimura KD, Seki T, Sato C, Ohno N, Sawamoto K. Neuraminidase inhibition promotes the collective migration of neurons and recovery of brain function. EMBO Mol Med, 16(6), 1228–1253 (2024).
17) Paredes MF, James D, Gil-Perotin S, Kim H, Cotter JA, Ng C, Sandoval K, Rowitch DH, Xu D, McQuillen PS, Garcia-Verdugo JM, Huang EJ, Alvarez-Buylla A. Extensive migration of young neurons into the infant human frontal lobe. Science, 354(6308), aaf7073 (2016).
18) Martí-Fàbregas J, Romaguera-Ros M, Gomez-Pinedo U, Martínez-Ramírez S, Jimenez-Xarrie E, Marin R, Martí-Vilalta JL, García-Verdugo JM. Proliferation in the human ipsilateral subventricular zone after ischemic stroke. Neurology, 74(5), 357–365 (2010).
19) Jin K, Wang X, Xie L, Mao XO, Zhu W, Wang Y, Shen J, Mao Y, Banwait S, Greenberg DA. Evidence for stroke-induced neurogenesis in the human brain. Proc Natl Acad Sci USA, 103(35), 13198–13202 (2006).

This page was created on 2025-07-11T14:07:28.023+09:00
This page was last modified on 2025-10-23T08:26:18.000+09:00
このサイトは(株)国際文献社によって運用されています。